β-lactamase enzymes have generated significant interest due to their ability to confer resistance to the most commonly used family of antibiotics in human medicine. Among these enzymes, the class B β-lactamases are members of a superfamily of metallo-β-lactamase (MβL) fold proteins which are characterised by conserved motifs (i.e., HxHxDH) and are not only limited to bacteria.
- metallo-β-lactamase (MβL) fold proteins
- multifunctional enzymes
- antibiotic-hydrolysing activity
1. Introduction
2. Diversity of the Superfamily of Metallo-β-Lactamase (MβL) Fold Enzymes
Within the classification of bacterial β-lactamases, which consist of four classes: A, B, C, and D [6][7][7,8], MβL enzymes alone occupy class B which is subdivided into three subclasses (subclass B1/B2/B3) as the result of divergent evolution events. They differ from the other classes by their active site which requires bivalent metal ions (such as Zn2+, Fe2+, Mg2+, or Ca2+) for their activity [6][8][9][10][7,9,10,11]. This catalytic site, characterised by a highly conserved motif (HxHxDH) and residues (H196 and H263), appears to be ancestrally shared by the superfamily of MβL fold proteins (with 34,000 proteins identified to date) with diverse biological functions including β-lactamases, nucleases, ribonucleases, lactonases, glyoxalases, hydrolases, phosphodiesterases, Aryl sulfatases, Alkylsulfatases, CMP-NeuAc hydroxylases, flavoproteins, and others which are distributed across the different domains of life including bacteria, archaea, and eukaryotes [11][12][13][12,13,14]. Moreover, as reported, the multifunctionality of these MβL fold enzymes and their affinities to different substrates are supported by the protein variable region of the enzymes (rather than the conserved motif (i.e., HxHxDH) and cofactor dependencies, which are responsible for the modulation of enzymatic activity, specificity, and oligomerisation proteins [9][14][10,15]. However, the protein similarity between enzymes of this MβL fold superfamily can be less than 20% [12][15][13,16] making it very unlikely that homologous sequences of contemporary bacterial MβLs in the other microorganisms such as Archaea, Giant viruses, Asgard, Nanoarchaeota, or Candidate Phyla Radiation (CPR) will be found. As reported in the “Darwinian Grandparenting” theory, we are genetically closer to our grandparents than to our cousins [16][17]; thus, the use of inferred common ancestor sequences as queries to identify homologous bacterial MβLs from not-yet investigated microorganisms represents one of the more pertinent approaches to identifying these enzymes in any domain of life. More sensitive approaches, including reconstruction of the common ancestor sequence [5][17][5,18], searching for Hidden Markov Models (HMM) profiles [17][18][18,19], Sequence Similarity Network analysis (SSN) [12][19][13,20], and 3D structure similarity analysis, have been used to look for homologous MβL enzymes in remote sources. As expected, these approaches appear significantly more flexible than classic Blast analysis in terms of identifying homologous MβL enzymes in unsuspected organisms and/or microorganisms.3. MβL Fold Enzymes in Bacteria: Class B β-Lactamases
3.1. Distribution and Diversity of MβL Fold Enzymes in Bacteria
In bacteria, these MβL enzymes include more than 325 variants grouped into 63 MβL types and divided into three sub-groups: subgroup B1 (e.g., NDM-1, VIM-2, and IMP-1); sub-group B2 (e.g., CphA1, CphA7, and ImiS); sub-group B3 (e.g., GOB-13, LRA-1, and CAR-1) (Figure 1), based on their differences in amino acid sequences and their catalytic sites which interact with either one or two Zn2+ ions [20][21][22][21,22,23]. The metallo-β-lactamase enzyme was identified for the first time in 1966, in a non-pathogenic strain of Bacillus cereus with a cephalosporinase activity which was inhibited when pre-incubated with ethylene-diamine-tetra-acetic acid (EDTA) [23][24]. Nowadays, MβL enzymes can be identified in more than 50 different bacterial species including gram-negative bacteria such as Enterobacteriaceae spp. and non-fermentative bacteria (such as, Acinetobacter spp. and Pseudomonas spp.), Elizabethkingia spp., Stenotrophomonas spp., Neisseria spp., Aeromonas spp., Shewanella spp., Myroides spp., Pedobacter spp., Empedobacter spp., Bacteroides spp., Vibrio spp., Bradyrhizobium spp., Caulobacter spp., Eristalis spp., Sphingomonas spp., Massilia spp., Burkholderia spp., Pectobacterium spp., and Gemmatimona spp.) [20][21][22][21,22,23]. Interestingly, as shown in Figure 1, while all the bacterial B1 and B2 MβL enzymes group together separately to the other domains of life, the GOB type enzymes (sub-group B3), are strongly related with enzymes from archaea and archaea-related microorganisms (i.e., Asgard), suggestive of their origin from this domain of life, and a horizontal transfer has occurred from archaea to a single bacterial group, namely, the Flavobacteriacaea family, especially in Elizabethkingia species. All the bacterial MβLs presented in this phylogenetic tree have been reported with proof of hydrolase activity on β-lactams, especially on carbapenems, and are all described in gram-negative bacteria except one MβL type i.e., BcII MβL enzyme (with seven variants), reported in a single bacterial gram-positive species, Bacillus cereus [20][24][21,25]. However, based on the MβL signature, these proteins have also been identified in other Gram-positive bacteria such as Streptococcus pneumoniae, in which MβL fold proteins are recognised as choline-binding proteins, DNA uptake-related proteins (nucleases) and L-ascorbate 6-phosphate lactonase [25][26][27][26,27,28]. Interestingly, it has been reported that the enzymatic characterisation of the L-ascorbate 6-phosphate lactonase enzyme from S. pneumoniae ATCC 49136 shows a β-lactamase activity since the purified enzyme is able to hydrolyse both nitrocefin and ampicillin-based antibiotics [26][27].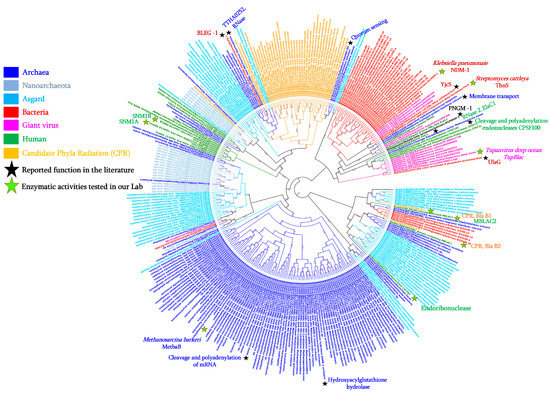
3.2. Reported Activities of Bacterial MβL Enzymes Other Than β-Lactams Hydrolysis
Besides their hydrolytic activities on β-lactam antibiotics, some bacterial MβL enzymes have been reported with other enzymatic activities as a result of the striking similarity between their protein structures and other enzymes including ribonuclease, nuclease, and lactonase enzymes. Indeed, as reported recently, the classical bacterial MβL IMP-1 enzyme, which hydrolyses all β-lactams including carbapenems, exhibits significant protein structure similarity with tRNase Z, a tRNA 3′ processing endoribonuclease of the MβL superfamily from Thermotoga maritima. Its enzymatic characterisation demonstrates a significant RNA-hydrolysing activity on both cellular RNA and synthetic small unstructured RNAs [28][29]. Interestingly, while this study was being published, the ouresearchers' research was beginning to reveal the ribonuclease and nuclease ability of the IMP-1 homologous enzyme i.e., class B NDM-1, described in almost all gram-negative bacteria, which significantly hydrolyses in vitro bacterial RNA and single-strand DNA substrates (Supplementary Figure S1). Moreover, while both bacterial MβL enzymes mentioned above can interact with RNA and/or DNA, others such as the ThnS enzyme can exhibit additional activities, such as the hydrolysis of ascorbic acid, as a result of its similarity with UlaG enzymes [29][30]. Indeed, as thwe researchers reported recently, while the thnS gene, part of the thienamycin (now chemically modified into imipenem in human medicine) biosynthesis gene cluster from Streptomyces cattleya, is annotated as putative β-lactamase with no reported proof of this activity. ThWe researchers demonstrated its specific hydrolase activity and UlaG high affinity with imipenem in comparison with the other β-lactams (e.g., penicillin G and cefotaxime). As a result of the phylogenetic tree and conserved motif analyses, the ThnS enzyme appears to be a member of the superfamily of MβL fold enzymes, showing additional activities of ribonuclease, nuclease, and hydrolysis of ascorbic acid [29][30]. Recently, an MβL fold enzyme (BLEG-1) has been reported in the Bacillus lehensis G1 strain, exhibiting significant sequence similarity and activity with the B3 subclass of bacterial MβLs, despite its evolutionary divergence from them [30][31][31,32] (Table 1). Upon analysing the phylogenetic tree and comparing the protein structures, it was discovered that the enzyme possessed an active site that was remarkably similar to those found in both the L1 B3 MβL from Stenotrophomonas maltophilia and the glyoxalase II enzymes (YcbL and GloB) from Salmonella enterica. Interestingly, the enzymatic characterisation of the purified BLEG-1 protein demonstrates its dual β-lactams hydrolysis (e.g., ampicillin hydrolysis) and glyoxalase activities [30][31]. The authors identify an insertion of two amino acids into the active-site loop at the N-terminal region of the BLEG-1 protein and suggested an evolution of the BLEG-1 enzyme from glyoxalase II to the adopted MβL fold activity through this insertion of amino acids [30][31].|
Kingdom |
Acc. Number |
Species |
Gene Name |
Size (aa) |
Default Annotation |
Reported Activities |
References |
|---|---|---|---|---|---|---|---|
|
Bacteria |
AGM20433 |
Enterobacter cloacae |
IMP-1 |
247 |
Imipenem hydrolysing β-lactamase |
β-lactamase; Ribonuclease |
|
|
AEW99090 |
Streptomyces cattleya |
ThnS |
330 |
Putative β-lactamase |
Imipenemase; Ascorbic acid degradation; Nuclease; Ribonuclease |
||
|
HQ328085 |
Klebsiella pneumoniae |
NDM-1 |
270 |
New Delhi metallo-β-lactamase |
β-lactamase; Ribonuclease |
In this study |
|
|
NA |
Streptococcus pneumoniae |
- |
363 |
L-ascorbate 6-phosphate lactonase |
β-lactamase |
||
|
P39300.2 |
Escherichia coli |
UlaG |
354 |
L-ascorbate-6-phosphate lactonase |
β-lactamase; Ribonuclease |
||
|
WP_010974862 |
Agrobacterium tumefaciens |
AiiB |
276 |
Zn-dependent hydrolases |
Quorum-quenching lactonase |
||
|
PDB: 7EV5_A |
Bacillus lehensis |
BLEG-1 |
210 |
β-lactamase domain containing protein |
β-lactamase; Glyoxalase II |
||
|
NA |
Soil metagenome |
MβLp01 |
312 |
Metallo-β-lactamase fold protein |
β-lactamase; Phytase |
||
|
NA |
Soil metagenome |
MβLP02 |
355 |
Metallo-β-lactamase fold protein |
β-lactamase; Phytase |
||
|
PDB: 7BZ4_B |
Deep-seep sediments metagenome |
PNGM-1 |
372 |
Metallo-β-lactamase |
β-lactamase; Ribonuclease |
||
|
WP_063100708 |
Escherichia coli |
YjcS |
661 |
Uncharacterised protein |
Alkyl sulfatase |
||
|
Archaea |
ELY35639 |
Haloferax volcanii |
HVO_2763 |
278 |
Zn-dependent hydrolases of the β-lactamase fold |
Membrane transport; Static and dynamic osmo-response |
|
|
851225341 |
Methanosarcina barkeri |
MetbaB |
214 |
MβL fold metallo-hydrolase |
β-lactamase; Ribonuclease; and D-lactate hydrolase |
||
|
15623131 |
Sulfolobus tokodaii |
- |
200 |
Putative hydrolase |
Quorum sensing activity |
[38] |
|
|
AAM30391 |
Methanosarcina mazei |
- |
638 |
Metal-dependent RNase, contains metallo-β-lactamase and KH domains |
Cleavage and Polyadenylation of mRNA |
[39] |
|
|
BAD70075 |
Thermus thermophilus |
TTHA0252 |
Metallo-β-lactamase superfamily |
Ribonuclease (RNase) |
[40] |
||
|
NA |
Methanocaldococcus jannaschii |
MjRNase J1 |
Ribonuclease Rnase J |
Ribonuclease (RNase) |
[41] |
||
|
NA |
Methanocaldococcus jannaschii |
MjRNase J2 |
Ribonuclease Rnase J |
Ribonuclease (RNase); Nuclease |
|||
|
NA |
Methanocaldococcus jannaschii |
MjRNase J3 |
Ribonuclease Rnase J |
Ribonuclease (RNase); Nuclease |
|||
|
Human |
NP_981951 |
Homo sapiens |
MβLAC2 |
280 |
Metallo-β-lactamase domain |
Exosome biogenesis enzyme; β-lactamase |
|
|
Q6PJP8 |
Homo sapiens |
SNM1A |
365 |
DNA cross-link repair 1A |
Cisplatin or Mitomycin hydrolase; β-lactamase |
||
|
Q9H816 |
Homo sapiens |
SNM1B |
335 |
5′ exonuclease Apollo isoform X1 |
β-lactamase |
[43] |
|
|
Giant viruses |
AUL78925 |
Tupanvirus deep ocean |
TupBlac |
322 |
β-lactamase superfamily domain |
β-lactamase; Ribonuclease |
[45] |
|
Candidate Phyla Radiation (CPR) |
KKR15801 |
Candidatus Levybacteria |
- |
287 |
β-lactamase class A-like protein |
β-lactamase; Ribonuclease |
[46] |
|
KKR17584 |
Candidatus Levybacteria |
- |
303 |
β-lactamase class A-like protein |
β-lactamase; Ribonuclease |
||
|
OGK62163 |
Candidatus Roizmanbacteria |
- |
263 |
Hypothetical protein |
β-lactamase; Ribonuclease |
||
|
OGZ64179 |
Candidatus Staskawiczbacteria |
- |
251 |
Hypothetical protein |
β-lactamase; Ribonuclease |
||
|
QHU90009 |
Candidatus Saccharibacteria |
- |
722 |
RNase J family beta-CASP ribonuclease |
β-lactamase; Ribonuclease |