2. Lipid Transporters (ABCA1 and ABCC1)
Lipids are a diverse class of biomolecules. Glycerophospholipids, specifically phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylserine (PS), and phosphatidylinositol (PI), as well as sphingolipids and cholesterol, serve as building blocks for membranes and organelles
[35][51]. Some ATP-binding cassette (ABC) transporters (ABCC1
[36][37][52,53]) act as “floppases” by catalyzing the movement of specific phospholipid species from the cytosolic leaflet to the extracellular leaflet of the plasma membrane (PM)
[38][54], while others (ABCA1
[39][55]) function as extracellular phospholipid translocases (
Figure 1). Indeed, ABC transporters have been shown to contribute to the asymmetric distribution of different phospholipids in the lipid bilayer, with PC and sphingolipids such as sphingomyelin (SM) residing predominantly in the outer leaflet of the PM, whereas anionic lipids such as PE, PS, and PI accumulate in the inner leaflet
[40][41][56,57]. Increasing evidence indicates that changes in the composition and distribution of these phospholipids in the lipid bilayer can regulate signal transduction pathways that are known to regulate PSC cell fates
[42][43][58,59].
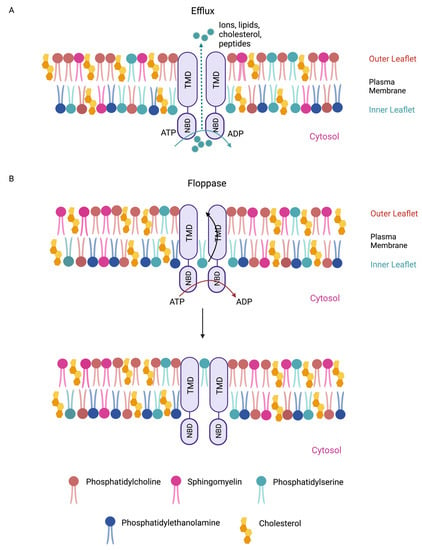
Figure 1. Membrane-bound mammalian ABC transporters are efflux pumps and/or floppases. (A) ABC proteins are essential membrane-bound transporters. ABC transporters are anchored at cell membranes through their transmembrane domains (TMDs). ABC transporters can efflux ions and macromolecules (e.g., lipids, cholesterol, and peptides) across cell membranes. (B) ABC transporters are also lipid floppases. They are critical for maintaining the asymmetric distribution of phospholipids in the membrane. Phosphatidylcholine (beige) and sphingomyelin (pink) reside predominantly in the outer leaflet of the plasma membrane, whereas anionic lipids such as phosphatidylserine (cyan) and phosphatidylethanolamine (blue) are more prevalent in the inner leaflet. Membrane cholesterols are depicted (yellow). Phospholipids such as phosphatidylserine from the inner membrane leaflet are “flopped” to the outer leaflet. Efflux and floppase activities require ATP hydrolysis by the nucleotide-binding domains (NBDs).
Stem cell maintenance in human PSCs requires basic fibroblast growth factor (bFGF), which activates the RAS-RAF-MEK-ERK signal transduction cascade
[44][45][46][47][60,61,62,63]. The association of RAS with the inner leaflet of the PM is an important step in the recruitment and activation of its effectors such as RAF and phosphatidylinositol 3-kinase (PI3K)
[48][64]. Interestingly, it has been shown that RAS can adopt a distinct orientation at the PM, depending on the types of phospholipids (PC, PS, or phosphatidylinositol 4,5-bisphosphate [PIP2]) that interact with RAS
[49][65]. As a result, the catalytic domain of membrane-bound RAS is predicted to become more exposed or partially obscured. Therefore, how RAS is anchored in the PM could modulate its ability to interact with its effectors (e.g., RAF versus PI3K) and regulate RAS-mediated downstream signaling choices. It appears that electrostatic interactions between RAS and lipids dictate interaction affinity and orientation preferences. Given that ABC transporters can translocate PC, PS, and PIP2 to the cell membrane outer leaflet
[50][51][52][66,67,68],
rwe
searchers propose that changes in the local distribution of phospholipids in the lipid bilayer by specific ABC transporters could influence the spatial arrangement of RAS. Future studies will be required to address the expression patterns of ABC transporters and their function in regulating the distribution of membrane phospholipids and RAS signal transduction, thereby controlling stem cell self-renewal versus differentiation. In a similar manner, it will be prudent to examine whether or not other signaling pathways (e.g., TGF-β
[53][69] and EGFR
[54][70]) that are known to contribute to stem cell pluripotency can also be modulated by PM phospholipid organization.
3. Cholesterol Transporters (ABCA1 and ABCG1)
Cholesterol is an important constituent of cell membranes. The bulk of cellular cholesterol (~90%) is localized at the PM
[55][71]. Cholesterol homeostasis is determined by the biosynthesis, uptake, and efflux of cholesterol. ABCA1 and ABCG1 play crucial roles in the efflux of cellular cholesterol and thus are important regulators of membrane cholesterol level
[56][57][58][72,73,74].
Cholesterol is a key modulator of membrane fluidity
[59][60][75,76], which in turn regulates cell behaviors such as adhesion, proliferation, and migration
[61][77]. However, recent evidence indicates that changes in PM stiffness may also regulate cell fate changes in PSCs
[62][78]. It has been shown that the rigidification of the PM precedes or coincides with downregulation of gene expression programs that stabilize the pluripotent state in PSCs, suggesting that a decrease in membrane fluidity may prime PSCs to exit from pluripotency. Consistent with the notion that maintenance of membrane fluidity contributes to stem cell maintenance, enzymes in the cholesterol biosynthesis pathways have been shown to be expressed at higher levels in PSCs, thereby increasing membrane cholesterol content and fluidity
[62][63][78,79]. Importantly, the inhibition of cholesterol production in PSCs accelerates their exit from pluripotency, as indicated by the rapid downregulation of stem cell marker alkaline phosphatase
[62][78]. These observations underscore the importance of cholesterol homeostasis in stem cell maintenance.
ResWe
archers propose that dissecting the mechanisms by which the expression and activities of ABCA1 and ABCG1 are controlled in PSCs will advance our understanding of the role of cholesterol efflux in regulating membrane fluidity and stem cell pluripotency.
In addition to regulating membrane fluidity, cholesterol, together with SM, has been shown to assemble dynamic, cholesterol-rich microdomains in the outer leaflet of the PM
[64][80]. These compartmentalized domains, known as lipid rafts, have been shown to enrich specific receptors and their effectors to promote receptor–effector interactions, thereby lowering activation barriers. The ability of lipid rafts to partition and concentrate select signaling machineries depends on the intrinsic affinity of these signaling proteins to lipid rafts, which has been shown to be influenced by amino acid sequences in the TM domains of membrane receptors and protein palmitoylation
[65][66][81,82]. Oligomerization of receptors has also been reported to increase their affinity to lipid rafts and residence time in these lipid subdomains
[67][83], hinting at a potential mechanism by which lipid rafts amplify signaling.
RWe
searchers suggest that a small change in the concentration of signaling components in lipid rafts may be sufficient, through amplification, to initiate signaling cascades. Therefore, lipid rafts may play an important role in increasing the responsiveness of signal transduction machineries to cellular stimuli.
It has been shown that ABCA1 and ABCG1 deficiency in macrophages leads to an increase in the number of lipid rafts and enhanced signaling responses
[68][84]. This is likely due to the propensity of lipid rafts to cluster, resulting in the amplification of signals
[69][70][85,86]. These observations suggest an inhibitory function of ABCA1 and ABCG1 in lipid raft formation, via the mobilization of cholesterol from lipid rafts to non-raft domains. It will be of interest to determine the mechanisms by which ABC transporters are recruited to lipid rafts. This is because the active efflux of membrane cholesterol by ABC transporters could facilitate the fine-tuning and dissolution of signal transduction hubs in lipid rafts and signal termination.
Lipid rafts are also detected in PSCs, but their roles in stem cell maintenance are less well-understood
[71][87]. The self-renewal of mouse PSCs requires leukemia inhibitory factor (LIF) signaling
[72][88]. It has been shown that depletion of membrane cholesterol in mouse PSCs by methyl-β-cyclodextrin (Mβ-CD), which has been shown to disrupt lipid rafts, compromises the recruitment of LIF receptor and its co-receptor gp130 to rafts and blunts LIF receptor-JAK-STAT3 signaling
[71][87]. The observed reduction in expression levels of key pluripotency-associated transcription factors OCT4 and SOX2 in Mβ-CD-treated PSCs indicates a destabilized pluripotent state when lipid raft formation is impaired. These observations are consistent with the role of lipid rafts in enriching specific receptors and facilitating their activation. Lipid rafts have also been implicated in other signaling pathways that are known to promote stem cell self-renewal and pluripotency, such as EGFR
[54][70] and RAS
[73][89], and those that destabilize the stem cell state, including insulin receptor
[74][90] and hedgehog
[75][91]. An outstanding question is how ABC transporters may control lipid raft formation and dynamics to partition competing signaling in PSCs to favor self-renewal over differentiation.
4. Redox Regulation and Oxidative Stress (ABCC1 and ABCC4)
ROS are natural byproducts of cellular metabolism. ROS can cause damage to the basic building blocks of cells including DNA, protein, and lipids. Therefore, ROS pose significant threats to the ability of PSCs to maintain genome and proteome integrity as they self-renew. In addition to cellular damages inflicted by ROS build-up, an imbalance in ROS levels can also lead to the misregulation of redox sensor molecules via the oxidation of cysteine residues. Some of these redox sensors are key signaling effectors such as AKT and MAPK
[76][77][92,93]. Therefore, it is conceivable that an increase in ROS concentration destabilizes the pluripotent cell state in part by interfering with signaling pathways essential for stem cell maintenance
[78][94]. ROS levels in cells are determined by the rate of ROS generation and the rate of ROS scavenging by antioxidants. PSCs are able to maintain relatively low ROS levels compared to those of differentiated cells, in part due to their reliance on glycolysis rather than oxidative phosphorylation for energy production, which is known to generate less ROS
[79][80][95,96]. Nevertheless, the neutralization of ROS species by antioxidants remains a critical mechanism in regulating ROS homeostasis in PSCs as it is essential for stem cell maintenance
[81][82][97,98].
Glutathione (GSH) is a major antioxidant in cells
[83][84][99,100]. GSH levels are balanced by its synthesis, transport, efflux, and degradation. Studies have shown that ABCC1 is a major GSH exporter and can regulate intracellular GSH levels. The overexpression of ABCC1 reduces intracellular GSH levels, while ABCC1 deficiency increases GSH concentrations
[85][86][101,102]. Importantly, ABCC1 can export both GSH and various oxidized glutathione derivatives (e.g., glutathione disulfide (GSSG)), although with distinct substrate affinity
[87][88][89][103,104,105]. Therefore, in addition to cellular enzymes that can degrade GSH (e.g., CHAC1
[90][91][106,107]) or regenerate GSH from GSSG (e.g., GSH reductase
[92][108]), ABCC1 likely plays an integral role in maintaining the redox equilibrium in PSCs. It has been shown that oxidative stress downregulates key PSC-specific transcription factors OCT4 and SOX2, and compromises AKT signaling
[81][97]. While the precise mechanism is unclear, the destabilization of OCT4 proteins and inactivation of AKT via the oxidation of critical cysteines residues could compromise the gene transcription and cellular signaling required for stem cell maintenance
[76][93][94][92,109,110].
Like oxidative stress, reductive stress induced by excessive levels of GSH can also impair PSC functions. Physiological levels of ROS have been shown to promote PSC proliferation and accurate DNA synthesis
[95][111]. High concentrations of antioxidants interfere with cell cycle progression and lead to the accumulation of DNA breaks
[96][112], likely due to the toxic effects of high antioxidant levels on the stability of cell cycle regulators and proteins involved in the DNA damage response and DNA repair
[95][111]. The balance between ROS and antioxidants must be optimal, as both extremes, oxidative and reductive stress, are damaging to PSCs. Functional studies on the role of ABCC1 and ABCC4 in PSCs will address the precise role of GSH/GSSG efflux in establishing a cellular redox state favorable for stem cell self-renewal and genome maintenance.