Atherosclerosis is a multifactorial process characterized by forming fibrofatty lesions within the arterial wall and is considered the leading cause of death worldwide. Improvement in treatment and prevention is crucial, especially in patients with T2D, a clinical syndrome expected to affect 783.2 million people by 2045. Therefore, the treatment guidelines for T2D patients recommend a patient-tailored approach based on lifestyle modifications and the choice of optimal therapeutic option. An ideal anti-diabetic drug should have the following characteristics: significant impact on weight and cardiovascular comorbidities, low risk of hypoglycemia and adverse events, and, last but not least, low costs. Even if no optimal medication exists, incretins represent one of the most attractive and promising options. The “incretin effect” indicates the amplification of pancreatic insulin secretion induced by these gastrointestinal tract-released hormones.
- atherosclerosis
- type 2 diabetes mellitus
- GLP-1 receptor agonists
1. Introduction
2. Physiological Mechanisms of Incretins
Atherosclerosis is a multifactorial process characterized by forming fibrofatty lesions within the arterial wall and is considered the leading cause of death worldwide [18][17]. Improvement in treatment and prevention is crucial, especially in patients with T2D, a clinical syndrome expected to affect 783.2 million people by 2045 [19][18]. Therefore, the treatment guidelines for T2D patients recommend a patient-tailored approach based on lifestyle modifications and the choice of optimal therapeutic option. An ideal anti-diabetic drug should have the following characteristics: significant impact on weight and cardiovascular comorbidities, low risk of hypoglycemia and adverse events, and, last but not least, low costs. Even if no optimal medication exists, incretins represent one of the most attractive and promising options [20][19]. The “incretin effect” indicates the amplification of pancreatic insulin secretion induced by these gastrointestinal tract-released hormones [21][20]. Incretins were demonstrated to reduce glucagon concentrations, improve insulin sensitivity, and slow down gastric filling in diabetic patients, with decreased free fatty acid concentrations and body weight. Moreover, beyond glycemic control, incretins protect the cardiovascular system [22][21]. The glucose-dependent insulinotropic polypeptide (GIP), a 42 amino acid hormone, and glucagon-like peptide-1 (GLP-1), a 31 amino acid hormone, are the most critical studied incretins. They bind to distinct G-protein-coupled receptors highly expressed on pancreatic β-cell surfaces. More specifically, while GLP-1 is secreted by L-cells in the ileum, colon, and rectum, GIP is released from K-cells located predominately in the duodenum and proximal gut after feeding. GIP circulates in 10-fold higher concentrations than GLP-1, whereas GLP-1 appears more potent than GIP. The two hormones are secreted in parallel but are not stored in the same cytoplasmic granules [23][22]. GIP and GLP1 can be degraded by DPP-4, an amino-peptidase transmembrane protein with a large extracellular domain and a flexible segment anchored in the cell membrane expressed in most cell types. The enzyme is responsible for cleaving and inactivating both two peptides at one of the last alanine residues [24][23]. Inhibition of DPP-4 and the use of injectable GLP-1RA are the two strategies for potentiate incretin receptor signaling [23][22]. However, studies on GIP monotherapy were unsuccessful; this could be explained by the fact that, in diabetic patients, the endocrine pancreas remains responsive to GLP-1, but it is no longer responsive to GIP, so this must represent the most likely reason for the reduced incretin action of this hormone [25][24].2.1. Acute and Chronic Effects of GLP-1 on Pancreatic β Cells
GLP-1 exerts acute and chronic functions on pancreatic cells by binding to its receptor (Figure 1). Acutely, it triggers most insulin release from these cells in a glucose-dependent manner. Hence, glucose enters the pancreatic β-cells through glucose transporter-2 (GLUT2). After phosphorylation, glycolysis, and the mitochondrial tricarboxylic acid (TCA) cycle, glucose determines adenosine triphosphate (ATP) production. Increased intracellular ATP concentration leads to K+ ATP-dependent channel closures with consequent accumulation of K+ ions and membrane depolarization. Thus, cell membrane depolarization causes voltage-dependent Ca2+ channel activation, the influx of Ca2+ ions, and exocytosis of a sub-pool of insulin granules, which contains ∼1–5% of available insulin. This initial rapid process is followed by the remaining entire insulin release mediated by GLP-1 binding to its receptor (GLP-1R), as previously mentioned, allowing the release of ∼95–99% of insulin granules [26][25]. GLP-1R is a G protein-coupled receptor whose activation induces a rapid increase in c-adenosine monophosphate (cAMP), protein kinase A (PKA), and exchange protein activated directly by cAMP (EPAC) upregulation and increased glucose-dependent insulin release [27][26]. Further to the stimulation of insulin secretion, the chronic effects of GLP-1RA on pancreatic β cells also consist of the deceleration of mass reduction.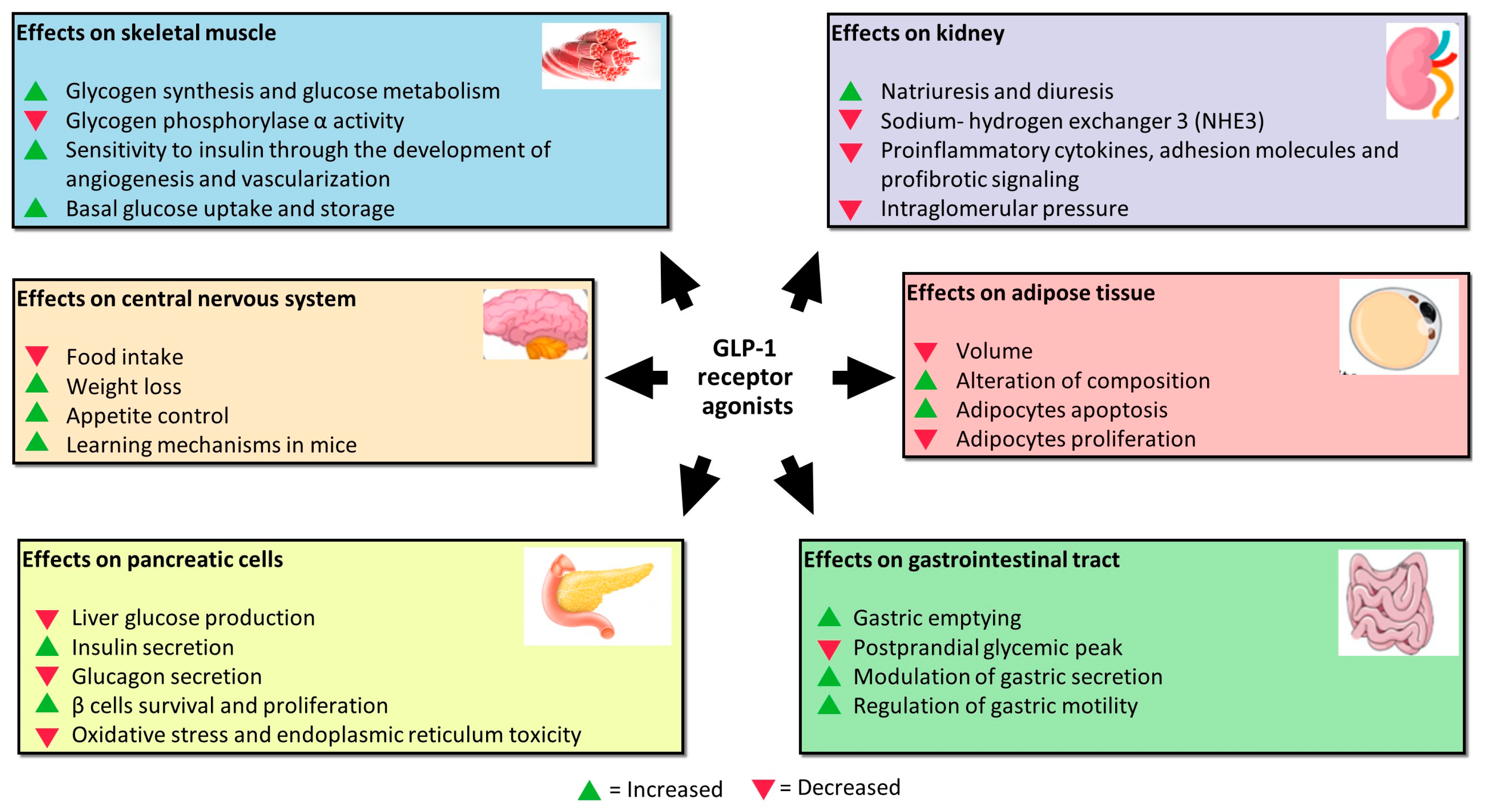
2.2. Effects of GLP-1 on Other Cell Lines
GLP-1 performs its function by acting not only on pancreatic β cells, but also on other cell lines (Figure 1). Studies assessing GLP-1RA extra-pancreatic effects reveal that exposure of skeletal muscle cells to these agents increases glycogen synthesis, glycogen synthase activity, and glucose metabolism and inhibits glycogen phosphorylase α activity involved in the breaking up of glycogen in glucose subunits [30][29]. Moreover, skeletal muscles exposed to increased incretin levels become more sensitive to insulin through more developed angiogenesis and vascularization. Indeed, GLP-1 binds to its receptor, abundantly expressed in endothelial cells, causing microvasculature recruitment in skeletal muscles and better muscle perfusion. Muscle perfusion is also favored via PKA-mediated endothelial nitric oxide synthase (eNOS) activation, which induces vasodilation and consequently increases insulin delivery [31][30]. These processes allow skeletal muscles to increase basal glucose uptake and storage. GlP-1 exerts a protective effect on the kidney. It is responsible for inhibiting sodium–hydrogen exchanger 3 (NHE3), an antiporter located on the epithelial cells of the proximal tube that imports sodium ions, simultaneously ejecting hydrogen ions in the proximal tubule lumen. NHE3 inhibition increases natriuresis and diuresis [32,33,34][31][32][33]. The mechanisms of this renal function gain may be direct and indirect. In addition to natriuresis, the first ones include the inhibition of proinflammatory cytokines, adhesion molecules, and profibrotic signaling, as well as the reduction in intraglomerular pressure through the inhibition of protein kinase-C (PKC) and the activation of PKA. The indirect mechanisms regard the benefit that GLP-1R agonists exert on other tissues, including improvement in blood pressure, glucose homeostasis, weight loss, and insulin levels that are beneficial for glomerular filtrate [35][34]. Furthermore, GLP-1 receptors were demonstrated to be located in many areas of the central nervous system; in particular, in those involved in appetite and gastric motility regulation [36][35]. GLP-1 is not only produced by alfa and L-cells, but also by neurons. It was proved that neuronally produced GLP-1 is transported to the axon terminals and stored in synaptic vesicles until release into the synaptic cleft, or in case of extra-synaptic release, into the brain parenchyma. Recent pre-clinical studies reported that administering GLP-1RA reduces food intake, determining weight loss in animal models [37][36]. The underlying mechanism is that L-cells-derived GLP-1 passes through the blood–brain barrier by directly affecting the receptors in the hypothalamic areas responsible for appetite control. GLP-1 is also released into the interstitial space near the site of its synthesis (ileum and colon) and then diffuses locally to act on vagal nerve endings embedded into the gut mucosa [38][37]. Moreover, the administration of GLP-1 in mice improves learning mechanisms, and the deletion of the gene encoding for GLP-1R is associated with neuron degeneration. Because of this, GLP-1RA was proposed as adjuvant therapy in the treatment of neurodegenerative pathologies such as Alzheimer with encouraging results [39][38]. In visceral adipose tissue, GLP-1 was demonstrated to reduce volume and alter the composition in vivo [40][39]. GLP-1 exerts this effect through activation of extracellular signal-regulated kinase (ERK), PKC, and AKT pathways with consequent increased adipocytes apoptosis and reduced pre-adipocytes proliferation [27][26]. After all, GLP-1 acts directly at the gastrointestinal level, inhibiting gastric emptying and postprandial glycaemic peak reduction due to the slower transit of food from the stomach to the small intestine. The mechanisms are still unknown, but GLP-1R is expressed on the parietal cells of the stomach to indicate a direct action of GLP-1 on their secretion. In addition, vagal denervation was shown to abolish the inhibitory effect of GLP-1 on gastric emptying, suggesting that GLP-1 acts through receptors expressed on vagal fibres that regulate gastric motility [41][40].References
- Pfeffer, M.A.; Claggett, B.; Diaz, R.; Dickstein, K.; Gerstein, H.C.; Køber, L.V.; Lawson, F.C.; Ping, L.; Wei, X.; Lewis, E.F.; et al. Lixisenatide in Patients with Type 2 Diabetes and Acute Coronary Syndrome. N. Engl. J. Med. 2015, 373, 2247–2257.
- Marso, S.P.; Daniels, G.H.; Brown-Frandsen, K.; Kristensen, P.; Mann, J.F.E.; Nauck, M.A.; Nissen, S.E.; Pocock, S.; Poulter, N.R.; Ravn, L.S.; et al. Liraglutide and Cardiovascular Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2016, 375, 311–322.
- Marso, S.P.; Bain, S.C.; Consoli, A.; Eliaschewitz, F.G.; Jódar, E.; Leiter, L.A.; Lingvay, I.; Rosenstock, J.; Seufert, J.; Warren, M.L.; et al. Semaglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N. Engl. J. Med. 2016, 375, 1834–1844.
- Holman, R.R.; Bethel, M.A.; Mentz, R.J.; Thompson, V.P.; Lokhnygina, Y.; Buse, J.B.; Chan, J.C.; Choi, J.; Gustavson, S.M.; Iqbal, N.; et al. Effects of Once-Weekly Exenatide on Cardiovascular Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2017, 377, 1228–1239.
- Hernandez, A.F.; Green, J.B.; Janmohamed, S.; D’Agostino, R.B.; Granger, C.B.; Jones, N.P.; Leiter, L.A.; Rosenberg, A.E.; Sigmon, K.N.; Somerville, M.C.; et al. Albiglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes and Cardiovascular Disease (Harmony Outcomes): A Double-Blind, Randomised Placebo-Controlled Trial. Lancet 2018, 392, 1519–1529.
- Gerstein, H.C.; Colhoun, H.M.; Dagenais, G.R.; Diaz, R.; Lakshmanan, M.; Pais, P.; Probstfield, J.; Riesmeyer, J.S.; Riddle, M.C.; Rydén, L.; et al. Dulaglutide and Cardiovascular Outcomes in Type 2 Diabetes (REWIND): A Double-Blind, Randomised Placebo-Controlled Trial. Lancet 2019, 394, 121–130.
- Husain, M.; Birkenfeld, A.L.; Donsmark, M.; Dungan, K.; Eliaschewitz, F.G.; Franco, D.R.; Jeppesen, O.K.; Lingvay, I.; Mosenzon, O.; Pedersen, S.D.; et al. Oral Semaglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N. Engl. J. Med. 2019, 381, 841–851.
- Gerstein, H.C.; Sattar, N.; Rosenstock, J.; Ramasundarahettige, C.; Pratley, R.; Lopes, R.D.; Lam, C.S.P.; Khurmi, N.S.; Heenan, L.; Del Prato, S.; et al. Cardiovascular and Renal Outcomes with Efpeglenatide in Type 2 Diabetes. N. Engl. J. Med. 2021, 385, 896–907.
- White, W.B.; Cannon, C.P.; Heller, S.R.; Nissen, S.E.; Bergenstal, R.M.; Bakris, G.L.; Perez, A.T.; Fleck, P.R.; Mehta, C.R.; Kupfer, S.; et al. Alogliptin after Acute Coronary Syndrome in Patients with Type 2 Diabetes. N. Engl. J. Med. 2013, 369, 1327–1335.
- Rosenstock, J.; Perkovic, V.; Johansen, O.E.; Cooper, M.E.; Kahn, S.E.; Marx, N.; Alexander, J.H.; Pencina, M.; Toto, R.D.; Wanner, C.; et al. Effect of Linagliptin vs Placebo on Major Cardiovascular Events in Adults with Type 2 Diabetes and High Cardiovascular and Renal Risk. JAMA 2019, 321, 69.
- Scirica, B.M.; Bhatt, D.L.; Braunwald, E.; Steg, P.G.; Davidson, J.; Hirshberg, B.; Ohman, P.; Frederich, R.; Wiviott, S.D.; Hoffman, E.B.; et al. Saxagliptin and Cardiovascular Outcomes in Patients with Type 2 Diabetes Mellitus. N. Engl. J. Med. 2013, 369, 1317–1326.
- Green, J.B.; Bethel, M.A.; Armstrong, P.W.; Buse, J.B.; Engel, S.S.; Garg, J.; Josse, R.; Kaufman, K.D.; Koglin, J.; Korn, S.; et al. Effect of Sitagliptin on Cardiovascular Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 232–242.
- Rosenstock, J.; Kahn, S.E.; Johansen, O.E.; Zinman, B.; Espeland, M.A.; Woerle, H.J.; Pfarr, E.; Keller, A.; Mattheus, M.; Baanstra, D.; et al. Effect of Linagliptin vs Glimepiride on Major Adverse Cardiovascular Outcomes in Patients with Type 2 Diabetes. JAMA 2019, 322, 1155.
- Visseren, F.L.J.; Mach, F.; Smulders, Y.M.; Carballo, D.; Koskinas, K.C.; Bäck, M.; Benetos, A.; Biffi, A.; Boavida, J.-M.; Capodanno, D.; et al. 2021 ESC Guidelines on Cardiovascular Disease Prevention in Clinical Practice. Eur. Heart J. 2021, 42, 3227–3337.
- Cosentino, F.; Grant, P.J.; Aboyans, V.; Bailey, C.J.; Ceriello, A.; Delgado, V.; Federici, M.; Filippatos, G.; Grobbee, D.E.; Hansen, T.B.; et al. 2019 ESC Guidelines on Diabetes, Pre-Diabetes, and Cardiovascular Diseases Developed in Collaboration with the EASD. Eur. Heart J. 2020, 41, 255–323.
- Brown, E.; Heerspink, H.J.L.; Cuthbertson, D.J.; Wilding, J.P.H. SGLT2 Inhibitors and GLP-1 Receptor Agonists: Established and Emerging Indications. Lancet 2021, 398, 262–276.
- Stanciulescu, L.A.; Scafa-Udriste, A.; Dorobantu, M. Exploring the Association between Low-Density Lipoprotein Subfractions and Major Adverse Cardiovascular Outcomes—A Comprehensive Review. Int. J. Mol. Sci. 2023, 24, 6669.
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, Regional and Country-Level Diabetes Prevalence Estimates for 2021 and Projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119.
- Drucker, D.J.; Holst, J.J. The Expanding Incretin Universe: From Basic Biology to Clinical Translation. Diabetologia 2023, 28, 1–5.
- Holst, J.J. The Incretin System in Healthy Humans: The Role of GIP and GLP-1. Metabolism 2019, 96, 46–55.
- Pahud de Mortanges, A.; Sinaci, E.; Salvador, D.; Bally, L.; Muka, T.; Wilhelm, M.; Bano, A. GLP-1 Receptor Agonists and Coronary Arteries: From Mechanisms to Events. Front. Pharmacol. 2022, 13, 856111.
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018, 27, 740–756.
- Gallwitz, B. Clinical Use of DPP-4 Inhibitors. Front. Endocrinol. 2019, 10, 389.
- Nauck, M.A.; Meier, J.J. The Incretin Effect in Healthy Individuals and Those with Type 2 Diabetes: Physiology, Pathophysiology, and Response to Therapeutic Interventions. Lancet Diabetes Endocrinol. 2016, 4, 525–536.
- Barg, S.; Huang, P.; Eliasson, L.; Nelson, D.J.; Obermüller, S.; Rorsman, P.; Thévenod, F.; Renström, E. Priming of Insulin Granules for Exocytosis by Granular Cl—Uptake and Acidification. J. Cell Sci. 2001, 114, 2145–2154.
- Rowlands, J.; Heng, J.; Newsholme, P.; Carlessi, R. Pleiotropic Effects of GLP-1 and Analogs on Cell Signaling, Metabolism, and Function. Front. Endocrinol. 2018, 9, 672.
- Wang, Q.; Brubaker, P. Glucagon-like Peptide-1 Treatment Delays the Onset of Diabetes in 8 Week-Old Db/Db Mice. Diabetologia 2002, 45, 1263–1273.
- Cornu, M.; Modi, H.; Kawamori, D.; Kulkarni, R.N.; Joffraud, M.; Thorens, B. Glucagon-like Peptide-1 Increases β-Cell Glucose Competence and Proliferation by Translational Induction of Insulin-like Growth Factor-1 Receptor Expression. J. Biol. Chem. 2010, 285, 10538–10545.
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like Peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130.
- Chai, W.; Dong, Z.; Wang, N.; Wang, W.; Tao, L.; Cao, W.; Liu, Z. Glucagon-Like Peptide 1 Recruits Microvasculature and Increases Glucose Use in Muscle via a Nitric Oxide–Dependent Mechanism. Diabetes 2012, 61, 888–896.
- Pyke, C.; Heller, R.S.; Kirk, R.K.; Ørskov, C.; Reedtz-Runge, S.; Kaastrup, P.; Hvelplund, A.; Bardram, L.; Calatayud, D.; Knudsen, L.B. GLP-1 Receptor Localization in Monkey and Human Tissue: Novel Distribution Revealed with Extensively Validated Monoclonal Antibody. Endocrinology 2014, 155, 1280–1290.
- Nusca, A.; Piccirillo, F.; Viscusi, M.M.; Giannone, S.; Mangiacapra, F.; Melfi, R.; Ricottini, E.; Ussia, G.P.; Grigioni, F. Contrast-Induced Acute Kidney Injury in Diabetic Patients and SGLT-2 Inhibitors: A Preventive Opportunity or Promoting Element? J. Cardiovasc. Pharmacol. 2022, 80, 661–671.
- Gutzwiller, J.-P.; Tschopp, S.; Bock, A.; Zehnder, C.E.; Huber, A.R.; Kreyenbuehl, M.; Gutmann, H.; Drewe, J.; Henzen, C.; Goeke, B.; et al. Glucagon-Like Peptide 1 Induces Natriuresis in Healthy Subjects and in Insulin-Resistant Obese Men. J. Clin. Endocrinol. Metab. 2004, 89, 3055–3061.
- Skov, J. Effects of GLP-1 in the Kidney. Rev. Endocr. Metab. Disord. 2014, 15, 197–207.
- Jessen, L.; Smith, E.P.; Ulrich-Lai, Y.; Herman, J.P.; Seeley, R.J.; Sandoval, D.; D’Alessio, D. Central Nervous System GLP-1 Receptors Regulate Islet Hormone Secretion and Glucose Homeostasis in Male Rats. Endocrinology 2017, 158, 2124–2133.
- Ard, J.; Fitch, A.; Fruh, S.; Herman, L. Weight Loss and Maintenance Related to the Mechanism of Action of Glucagon-Like Peptide 1 Receptor Agonists. Adv. Ther. 2021, 38, 2821–2839.
- During, M.J.; Cao, L.; Zuzga, D.S.; Francis, J.S.; Fitzsimons, H.L.; Jiao, X.; Bland, R.J.; Klugmann, M.; Banks, W.A.; Drucker, D.J.; et al. Glucagon-like Peptide-1 Receptor Is Involved in Learning and Neuroprotection. Nat. Med. 2003, 9, 1173–1179.
- Perry, T.; Greig, N. A New Alzheimers Disease Interventive Strategy: GLP-1. Curr. Drug Targets 2004, 5, 565–571.
- Du, X.; Lu, W.; Lu, Z.; Shao, X.; Hu, C.; Shi, B. Exenatide with Metformin Ameliorated Visceral Adiposity and Insulin Resistance. J. Diabetes Res. 2018, 2018, 4019248.
- Baggio, L.L.; Huang, Q.; Brown, T.J.; Drucker, D.J. A Recombinant Human Glucagon-Like Peptide (GLP)-1–Albumin Protein (Albugon) Mimics Peptidergic Activation of GLP-1 Receptor–Dependent Pathways Coupled with Satiety, Gastrointestinal Motility, and Glucose Homeostasis. Diabetes 2004, 53, 2492–2500.