1. Introduction
Noncommunicable diseases (NCD) are disorders that are not transmissible from one person to another (e.g., obesity, diabetes, cancer, autoimmune diseases, cardiovascular diseases, respiratory, and musculoskeletal disorders) and are the result of a multifactorial combination of unhealthy lifestyle habits and genetic predisposition
[1]. They are among the leading causes of death, representing around 70% of total global deaths, and are recognized as one of the biggest challenges by the World Health Organization (WHO) and the Food and Agriculture Organization of the United Nations (FAO)
[2][3][2,3]. One of the leading causes of NCDs is malnutrition, which results from states of obesity, other dietary factors, and undernutrition (e.g., underweight, deficiencies in vitamins, stunting). On the other hand, ageing impacts biological and physiological functions, which promotes and aggravates the development of age-associated diseases (AAD) such as atherosclerosis, hypertension, cardiovascular diseases, Alzheimer’s disease, dementia, arthritis, and osteoporosis
[4]. Malnutrition also contributes to the onset of AADs and is often underdiagnosed in elderly patients
[5]. Therefore, intervention in dietary habits is a critical approach to tackle these challenges. Another issue associated with unhealthy dietary practices arises from the overexploitation and consequent pollution associated, for example, with current food production, which accounts for 20–35% global greenhouse emissions
[3]. A dietary shift to intake small amounts of calories from animal sources and increase the consumption of sustainable, nutrient-rich, and calorically efficient products, such as algae, are recommended by the FAO and WHO to prevent the aggravation of these issues
[3].
Most NCDs are associated with low-grade chronic inflammation, characterized by persistently elevated levels of circulating pro-inflammatory cytokines, chemokines, and acute inflammation phase proteins
[6]. The persistency of this low-grade chronic inflammation results in cellular, tissue, and organ damage over time, mainly through continuous oxidative stress, which eventually impairs proper body function. Mitigating the inflammatory response can prevent or decrease the severity of NCDs
[1]. On the other hand, AADs are associated with the increasing senescence processes associated with ageing, such as telomerase erosion, the oxidative damage of DNA and proteins, mitochondrial dysfunction, oncogene overexpression, or epigenetic factors, that result in stem cell exhaustion, the dysfunction of body systems, and chronic inflammation
[7]. To an extent, both NCD and AAD can be prevented by modifying lifestyle-related risk factors, where unhealthy diets play a significant role
[8][9][8,9]. The modulation of chronic inflammation through diet plays an important role in decreasing the risks and prevalence of NCDs and AADs, as diet can provide components such as omega-3 (ω-3) fatty acids, flavonoids, and vitamins, which can suppress inflammation, decrease oxidative damage, and modulate gene expression
[10].
The transition to more sustainable and healthy diets that are capable of preventing the development of NCDs and AADs, requires investment in and development of new food sources and ingredients with high nutritional value and diversity with sustainable production and reduced environmental impact (
Figure 1)
[3]. Microalgae are a sustainable and highly nutritious alternative that could enrich and contribute to the transition to more environmentally friendly and healthy diets
[11][12][11,12]. They are a diverse and rich source of nutritional components, such as vitamins (e.g., vitamin B12), carbohydrates, proteins, nucleic acids, lipids (e.g., ω-3 polyunsaturated fatty acids (PUFA), others), and functional components (e.g., chlorophylls, which have antioxidant activity)
[13]. Their lipid nutritional value is comparable to that of fish oils, offering an alternative to this commonly used source of ω-3 FA
[14][15][14,15]. Microalgae are considered a promising source of bioactive components, including bioactive lipids such as ω-3 lipids, essential precursors of anti-inflammatory eicosanoids, and polar lipids
[11]. Recent research suggests that marine polar lipids as phospholipids or glycolipids, which are highly concentrated in ω-3 PUFAs, could be more effectively delivered than triglycerides in terms of human health. Polar lipids are more stable and have higher bioavailability than triglycerides
[16]. Moreover, as a relevant fact, phospholipids have recently been suggested as an excellent vector of DHA and oxygenated DHA metabolites as protectins, enhancing their in vivo role as inflammation resolvers. Among these, glycolipids (GL) have shown chemotherapeutic potential, anti-proliferative effects, potent inhibition of nitric oxide (NO) release, and anti-inflammatory potential
[17]. Only a handful of microalgae species are approved for food consumption; however, the use of microalgae and their lipid extracts as food ingredients are recognized as having the potential to prevent NCDs and AADs and to enrich nutrient-deficient diets.
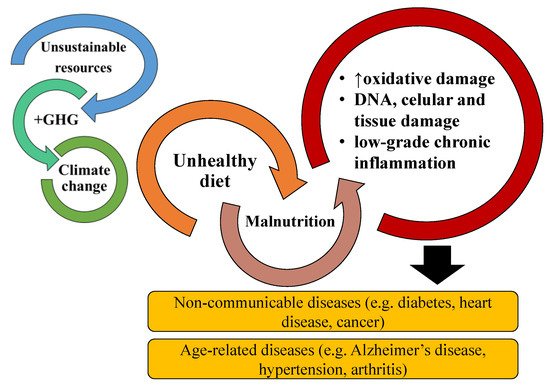
Figure 1. The relationship and risk of noncommunicable diseases and age-associated diseases are increased by malnutrition originating from poor and unhealthy diets. Malnutrition promotes increased oxidative stress, DNA, cellular and tissue damage, and chronic inflammation. Chronic inflammation is associated with most NCDs and AADs. Bad diets are also associated with the unsustainable exploitation of resources, greenhouse gases (GHG) emissions, and impacts on climate change.
On the other hand, the anti-inflammatory and antioxidant potential of microalgae lipid extracts were intensively studied in the past decade. In this context, this resviearchw will address the anti-inflammatory potential of bioactive raw extracts, lipidic fractions, and isolated lipids from microalgae. This information will help valorize and promote microalgal lipids as sustainable and healthy dietary approaches to tackle global malnourishment, NCD, AAD, and environmental degradation.
Literature Reviewing Strategy
This revision was performed using the platform Web of Science. The keywords used were a combination of “microalgae”, with “anti-inflammatory”, “immunomodulatory”, and immunomodulation. The inclusion criteria were the use of microalgae organic extracts or the use of microalgae isolated lipid classes and species that have been reported to have anti-inflammatory/pro-resolving effects. Out of 138 results, only 32 met the proposed criteria. The excluded works referred to the use of aqueous extracts. Other banned works involved evaluating the anti-inflammatory activity of pigments or other non-lipidic-based molecules that did not fit the purpose of this resviearchw.
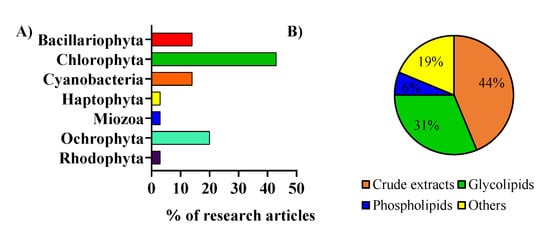
The included works were analyzed and were categorized by considering the microalga phylum (
Figure 2A) and the type of lipid extracts or fractions that were used (
Figure 2B). Microalgae from the phylum Chlorophyta were the most studied regarding anti-inflammatory activity. Most of the studies used crude lipid extracts in the anti-inflammatory activity assays
[18][19][20][18,19,20]. In contrast, others used lipid fractions in specific classes of lipids, such as glycolipids, phospholipids, or others (
Figure 2B)
[21][22][21,22]. Among these, the ones that used glycolipids from microalgae enriched fractions were the most studied
[23][24][25][23,24,25].
Figure 2. (A) Microalgae of different phyla were used in the assays to evaluate the anti-inflammatory activity of lipids. (B) Type of lipid extracts from microalgae used to assay the anti-inflammatory activity.
2. Microalgae Lipids: Structural Diversity and Functionality
The microalgae lipidome includes two main groups of lipids: neutral lipids (fatty acids, TAG, and sterols) and polar lipids (sphingolipids, phospholipids (PL), glycolipids (GL), and betaine lipids (BL)). Polar lipids can develop several biological functions, acting as the structural components of cell membranes, which constitute lipoproteins, energy reserves, and signaling molecules
[26].
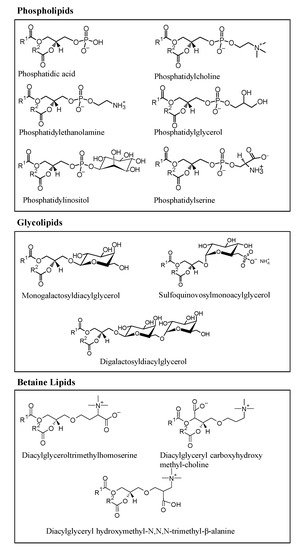
Microalgae are rich in polar lipids, constituting 41–92% of total lipids, while neutral lipids constitute 5–51%. The lipidomic profile of several microalgae species have been studied, and significant polar lipids, represented in
Figure 3, that have been identified include species from several classes of phospholipids, such as phosphatidic acid (PA), phosphatidylserine (PS), phosphatidylethanolamine (PE), phosphatidylinositol (PI), phosphatidylcholine (PC), phosphatidylglycerol (PG); glycolipids, such as digalactosyldiacylglycerol (DGDG), sulfoquinovosylmonoacylglycerol (SQDG), monogalactosyldiacylglycerol (MGDG); and betaines lipids, such as diacylglyceroltrimethylhomoserine (DGTS), diacylglyceryl hydroxymethyl-N,N,N-trimethyl-β-alanine (DGTA), and diacylglyceryl carboxyhydroxy methyl-choline (DGCC), as represented in
Figure 3. Some of the polar lipid species described here were reported as being biologically active
[27][28][29][30][31][32][33][27,28,29,30,31,32,33].
Figure 3. Main classes of polar lipids found in microalgae: glycerophospholipids (or phospholipids), glycoglycerolipids (or glycolipids), and betaine lipids.
Polar lipids and neutral lipids are the leading FA carriers, which are the most studied lipids in microalgae.
[34][35][36][37][38][34,35,36,37,38]. Microalgae FAs are typically 12 to 22 carbons long with up to 6 unsaturations; however, short-chain FA and oxidized PUFAs have been reported
[39][40][39,40]. FAs such as the omega-6 and arachidonic acid (ARA) and the ω-3 FAs, such as α-linolenic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA), are usually associated with the nutritional value and bioactive properties from algae. They are precursors of pro- and anti-inflammatory eicosanoids, respectively, and contribute to the balance of both omega-6 and ω-3 FA, which are important for the normal functioning of the immune system
[41][42][43][41,42,43].
Polar lipids have been described as potent bioactive compounds
[44][45][44,45], and therefore, the interest in understanding their bioactive mechanisms and the extent of their bioactive potential have increased. Several studies have pointed to PL and GL, especially those esterified with ω-3 PUFAs, as possessing antioxidant, anti-inflammatory, anti-obesity, anti-tumor, anti-viral and anti-bacterial activity
[16][17][46][47][16,17,46,47]. The bioactivities observed for these lipids hold exciting potential for natural sources prospection and can pose an alternative to the current sources of these lipids (e.g., fish oils). Fish oil is a commercially available ingredient and nutritional supplement rich in ω-3 PUFAs
[3][48][49][3,48,49]. Still, microalgae are considered a sustainable alternative to fish oils and as a source of healthy and bioactive ω-3 PUFAs.
Anti-inflammatory activity was one of the main biological activities reported for microalgae lipid extracts
[17]. Moreover, some microalgae polar lipids have been reported to have anti-inflammatory activity
[17][22][50][17,22,50]. The following section will discuss the anti-inflammatory activity observed for microalgae lipid extracts and fractionated lipids. These studies evaluated pro-inflammatory markers, and some tried to understand a possible relationship between the selected microalgae species, extracts, and fractions and their immunomodulatory activity. Distinct parameters such as the induction or attenuation of cytokine production, the gene expression of inflammatory markers, and the activation or inhibition of signaling pathways were approached.
3. The Anti-Inflammatory Potential of Microalgal Lipid Extracts
The anti-inflammatory properties of bioactive lipids were evaluated using crude extracts from several microalgae, namely Chlorella vulgaris, Chlorella ovalis, Nannochloropsis oculata, Nannochloropsis granulata, Nannochloropsis oceanica, Phaeoductylum tricornutum, Amphidinium carterae; the diatoms Odontella mobiliensis, Pseudonitzschia pseudodelicatissima, Coscinodiscus actinocyclus, and Alexandrium minutum; and the mutant microalgae species Tetraselmis sp. (IMP3 and CTP4) or cyanobacteria Arthrospira maxima
[19][32][33][50][51][52][53][54][55][19,32,33,50,51,52,53,54,55]. Different solvents and solvent systems were used to extract the lipids, making an accurate comparison of the results difficult. In these studies, the anti-inflammatory potential was accessed through the evaluation of the release/production of inflammatory mediators, such as the tumor necrosis factor α (TNF-α) and interleukin-6 (IL-6), prostaglandin E2 (PGE2) and NO, and the expression of the key enzymes, such as cyclooxygenase-2 (COX-2) and iNOS.
Very few studies were performed in animal models
[23][24][56][57][23,24,56,57]. The reduction of croton-induced oedema in mice and neutrophils concentration in the wound region of zebrafish was observed when treated with lipids from microalgae
[23][24][23,24]. Other studies observed the response of pro-inflammatory markers, namely cytokines, to microalgae extracted lipids in inflammatory disease models of induced colitis in rats and diabetic mice
[56][57][56,57]. The lack of studies performed in animal models of disease hampers elucidation of the complete impact of lipids in the complex network of inflammation.
3.1. In Vitro Evaluation of Microalgal Lipids Impact in Key Pro-Inflammatory Enzymes
The evaluation of COX-2 and iNOS activity and expression are the most common parameters measured in screenings for anti-inflammatory potential, both generally and for microalgal extracts. COX-2 is an enzyme induced by pro-inflammatory mediators that converts arachidonic acid to prostaglandins. The modulation of its activity and expression provides information regarding the production of pro-inflammatory prostaglandins with a critical role in the initial onset of inflammation
[58]. COX-2 inhibition was reported in studies using lipid extracts from
Gloeothece sp.,
Chlorella vulgaris grown under auto- and heterotrophic conditions,
Chlorococcum amblystomatis and lipid extracts enriched in ω-3 PUFAs from
Tetraselmis sp. mutant strains (IMP3 and CTP4)
, Skeletonema sp., and
Nitzschia palea [19][53][59][60][61][19,53,59,60,61]. These studies evaluated the inhibitory COX-2 activity of the crude lipids with a commercial assay in chemico
, as described in Supplementary Table S1. The evaluation of the capacity of lipid extracts to modulate COX-2 activity was also performed in in vitro cells studies, using lipid extracts from
Chlorella vulgaris, Chloromonas reticulata, Micractinium sp.,
Nannochloropsis oculata, Nitzschia palea, and
Phaeodactylum tricornutum. The results showed COX-2 inhibition and the downregulation of COX-2 protein levels in Raw264.7 cells
[20][52][60][62][63][64][65][20,52,60,62,63,64,65].
On the other hand, assessing iNOS activity and expression provides information regarding the production of NO, a key molecule for induction and inflammation maintenance. The most common strategy to assess the impact of the tested lipid extracts on iNOS activity relies on evaluating their effects on LPS-triggered NO production by Raw 264.7 macrophages
(Supplementary Table S1). For instance,
Amphidinium carterae,
Chlorella sp.,
Chloromonas reticulata,
Micractinium sp.,
Nannochloropsis oculata,
Nitzschia palea, and
Phaeoductylum tricornutum lipid extracts were shown to reduce NO levels and iNOS expression induced by LPS in Raw264.7 cells
[20][51][52][60][62][63][64][65][66][20,51,52,60,62,63,64,65,66]. Moreover, MGDG and DGDG
[67], two classes of glycolipids, and DGTS
[22], one of the betaine lipids, extracted from
Nannochloropsis granulata, revealed intense NO inhibitory activity in Raw264.7 macrophages. Finally, rats supplemented with
Nannochloropsis oculata extracted glycolipid-rich oil revealed a significant reduction of NO production through the downregulation of iNOS
[68].
Abu-Serie et al.
[63] reported that extracts from
Chlorella vulgaris reduced NF-κB expression together with iNOS and COX-2 in an in vitro model of lipopolysaccharide-stimulated white blood cells. The NF-κB signaling pathway plays a central role in inflammation by regulating the transcription of pro-inflammatory genes such as COX-2, iNOS, and multiple pro-inflammatory cytokines
[69]. Therefore, the decreased expression of COX-2 and iNOS caused by microalgae lipid extracts could result from the downregulation of this particular pathway, opening new target possibilities that require further study.
3.2. In Vitro Evaluation of Microalgae Lipids Impact in Pro-Inflammatory Cytokines
Another essential step to determine the anti-inflammatory potential of microalgae lipid extracts is evaluating the production/release of pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, in activated immune cells. Pro-inflammatory cytokines are predominantly produced by activated immune cells (e.g., macrophages, monocytes, lymphocytes) and play an essential role in pro-inflammatory reactions
[70]. Their inhibition results in the attenuation of the inflammatory response, posing a decisive step towards inflammation resolution. The assessment of TNF-α, IL-1β, and IL-6 levels in cell lines (Raw264.7 and THP-1 macrophages) reported for lipid extracts from
Nitzschia palea,
Chlorella vulgaris,
Tetraselmis suecica,
Micractinium sp.,
Aurantiochytrium mangrovei,
Phaeodactylum tricornutum,
Chloromona reticulata, and
Spirulina maxima showed the capacity to downregulate their production
[20][60][62][63][64][65][66][71][20,60,62,63,64,65,66,71]. When compared to the use of crude extracts to evaluate the downregulatory effect of microalgae lipids on pro-inflammatory cytokines, the studies using isolated lipid classes and species are reduced. Oxylipins resulting from the enzymatical oxidation of PUFAs, isolated from
Chlamydomonas debaryana and
Nannochloropsis gaditana, and LPC(16:0), isolated from
Cylindrotheca Closterium, showed the capacity to downregulate LPS-triggered TNF-α production in THP-1 macrophages
[72][73][72,73]. The induction of IL-6 in LPS-activated Raw264.7 cells was decreased with free and esterified DGLA from a mutant strain of the microalga
Lobosphaera incisa P127
[74]. Another study observed a reduction in TNF-α, IL-6, and IL-1β expression in peritoneal blood mononuclear cells (PBMC) treated with ergosterol and 7-dehydroporiferasterol isolated from the microalga
Dunaliella tertiolecta [75]. This mix of phytosterols from the microalga
Dunaliella tertiolecta raised anti-inflammatory cytokine IL-10 levels, strengthening the anti-inflammatory potential. Ávila-Román et al. described newly isolated oxylipins from
Chlamydomonas debaryana derived from 16:4 and 18:4 fatty acids and used them as diet supplementation in a 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced colitis animal model, observing a downregulation of TNF-α. The most active oxylipin was a C-16 hydroxy acid
[72].