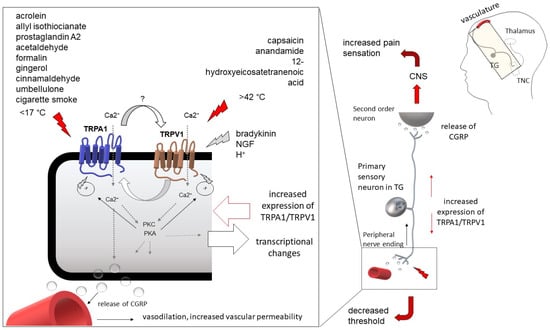
The transient receptor potential ankyrin 1 (TRPA1) has gained more attention in migraine-related research. The involvement of the TRPA1 receptor in migraine headaches is proposed by the fact that TRPA1 may be a target of some migraine-triggering factors. Although it is doubtful that activation of TRPA1 alone is sufficient to induce pain, behavioral studies have demonstrated that TRPA1 is involved in injury- and inflammation-induced hypersensitivity. Here, we review the functional relevance of TRPA1 in headaches and its therapeutic potential was introduced, mainly focusing on its role in the development of hypersensitivity, referring to its altered expression in pathological conditions, and its functional interaction with other TRP channels.
- TRPA1
- migraine
- hypersensitivity
1. Migraine
2. Transient Receptor Potential Ankyrin 1 (TRPA1)
In the last two decades, members of the TRP (transient receptor potential) superfamily have been identified to play a pivotal role in peripheral nociception and the sensation of pain [13][14][15][16][19,20,21,22]. TRPA1—formerly known as ankyrin-like with transmembrane domains protein 1 [ANKTM1], originally described by Story et al. [17][23]—is expressed by a small subpopulation of dorsal root ganglion neurons, as well as trigeminal neurons [18][19][24,25]. TRPA1 is activated by various noxious compounds such as allyl isothiocyanate (AITC), the pungent agent of mustard oil (MO), horseradish, and cinnamaldehyde [20][21][26,27]. Acrolein, an irritant compound found in tear gas and cigarette smoke, was the first substance to be identified as a ‘specific TRPA1 activator’, and, furthermore, electrophilic substances such as Δ9-tetrahydrocannabinol, icilin, and 2-aminoethoxydiphenyl borate (2APB) were also cited as TRPA1 activators [20][22][26,28]. The functional importance of the TRPA1 channel in peripheral pain sensation is suggested by the fact that TRPA1 expression is typically attributed to the small-sized neurons, commonly referred to as nociceptors. TRPA1-mediated pain sensation was reported in subjects following the administration of a synthetic TRPA1 agonist, suggesting that TRPA1 activation alone is sufficient to evoke pain in patients [23][29]. In addition, the importance of TRPA1 in human pain sensation is indicated by a genetic disorder in which TRPA1 is affected. Familial episodic pain syndrome, a genetic disorder accompanied by episodic body pain, has been shown to be associated with a TRPA1 deficiency [24][30].3. TRPA1 in Migraine-Related Research
3.1. The Presence of TRPA1 in the Trigeminal System
The migraine-inducing effect of environmental irritants has been associated with the activity of certain pain-related receptors over time. In recent decades, the classical TRP channel TRPV1, the mechanosensor Piezo channels, or the purinergic receptor PY have been identified as potential targets of certain migraine-triggering substances. In 2011, when Kunkler et al. reported TRPA1-mediated vasodilation in meningeal vessels, TRPA1 came into the focus of migraine-related research [25][47]. As TRPA1 has gained more attention in relation to head pain, the ratio of the TRPA1-expressing neurons in the intracranial sensory system involving the TG has become the subject of investigation. By using an in situ hybridization technique, it was found that TRPA1 mRNA is expressed in 36.5% of all trigeminal primary sensory neurons [26][48]. Similar results are obtained by functional studies, as the application of the TRPA1 agonist AITC stimulates nearly 35% of rat trigeminal neurons [20][26]. TG is responsible for the sensory innervation of the entire head region, including the facial skin, nasal mucosa, intracranial structures, meningeal vasculature, etc. Focusing on head pain, the neurons responsible for innervating the pain-sensitive structures within the skull must be distinguished from the overall population. Applying whole-cell patch-clamp recording, approximately 40% of the dural projection neurons responded to the application of the TRPA1 agonist MO, representing a slightly higher proportion than the total trigeminal neuronal population [27][49]. This result supports the substantial role of TRPA1 in intracranial sensory processes, as more than one-third of primary sensory neurons—considered potential pain-sensitive neurons—are involved in TRPA1-mediated reactions. Interestingly, only a small percentage of trigeminal neurons projecting to the dura show TRPA1-immunoreactivity [28][50]. However, the TRPA1-immunoreactive neurons cluster around most of the dural projection neurons, suggesting potential cross-excitation between the neighboring neurons.3.2. Relevance of TRPA1 in Migraine-Like Pain
NO was the first experimentally confirmed migraine-provoking factor, and its experimental administration has since been used as a widely accepted migraine model [29][58]. Tear gas was also considered a headache trigger, as in 29% of seizure events, patients complained about post-seizure headaches [30][59]. In recent decades, several natural compounds have been identified as headache-provoking factors: camphor, tear gas, formalin, cigarette smoke, or alcoholic beverages—all these agents are well-known headache triggers [31][60]. TRPA1 was originally identified as a ‘nociceptive’ receptor activated by thermal and mechanical stimuli; however, more and more headache-inducing factors have also been revealed to be TRPA1 activators. Several migraine triggers, such as formalin, cigarette smoke, and even tear gas, have been shown to act through the TRPA1 channel [25][32][33][34][47,61,62,63]. Moreover, the classical trigger, nitric oxide (NO), has also been found to induce pain sensations through the activation of TRPA1 [35][64]. In addition, an endogenous product, 4-hydroxynonenal, which is associated with several chronic diseases, has been shown to act as a potential activator of TRPA1, supporting the relevance of TRPA1-activation in chronic pain disorders such as migraine [36][65]. All of the above studies confirmed that certain migraine-inducing agents are potential activators of TRPA1; however, they did not unambiguously establish a direct link between the headache and TRPA1 receptor activation. It has recently been shown that TRPA1 contributes to migraine-associated hypersensitivity, since the TRPA1 antagonist, ADM_12, prevented the nitroglycerine-induced hyperalgesia in both the acute and chronic migraine models [37][46]. It was recognized early that headache attacks are accompanied by intracranial vascular reactions [38][39][66,67]. Vascular events were originally thought to cause the sensation of pain during headache attacks. However, the direct relationship between head pain and vascular reactions was disproved by modern imaging studies two decades ago [40][68]. At the same time, meningeal vasodilation is a clear consequence of the activation of chemosensitive trigeminal afferents, so the monitoring of intracranial vascular reactions following trigeminal nerve stimulation serves as an approved model for migraine. The phenomenon that inhalation of environmental irritants frequently triggers a migraine attack has led researchers to investigate the functional connection between nasal stimulation and headache-related meningeal events. The intranasal administration of chemical compounds and recording the evoked meningeal reactions provide a specific aspect of headache research. The putative role of TRPA1 in intracranial sensitization is supported by Kunkler et al. [25][47]. A nasal application of the TRPA1 activators MO and acrolein has been shown to increase the meningeal blood flow in a CGRP-dependent way. Subsequently, this group also demonstrated the sensitizing effect of TRPA1 activation on trigeminovascular responses, as chronic exposure to acrolein enhances TRPA1-mediated meningeal vasodilation and leads to facial allodynia [41][18]. In 2012, an experimental study showed that activation of TRPA1 via an injected dural cannula increased the pain sensation in the facial region [27][49]. Another behavioral study proved that umbellulone (UMB), the active compound of Umbellularia californica (‘headache tree’), causes trigeminal pain via TRPA1 [42][69]. It found that in 40% of rat trigeminal ganglion neurons, the administration of UMB could stimulate TRPA1, and intranasal and intravenous administration induced nociceptive behavior in a TRPA1-dependent manner4. Possible Cooperation between TRPA1 and TRPV1
It is well-documented that other classical TRP channels, e.g., the transient receptor potential vanilloid 1 (TRPV1), have a pivotal role in inflammation-related hypersensitivity such as thermal hyperalgesia and allodynia [43][44][45][72,73,74]. Hypersensitivity can develop as a result of a functional modification of the TRPV1 by phosphorylation-induced sensitization [46][47][48][75,76,77]. Considering the importance of intracranial sensitization in the pathomechanism of migraine, the functional sensitization of TRPA1, TRPV1, and other TRP receptors involved in intracranial pain sensation at the receptor level should be discussed as a provocative factor in migraine attacks. Based on its structure, TRPA1 belongs to the TRP superfamily. As a common feature, TRP subfamilies possess a tetrameric structure, each involving six transmembrane domains (S1 to S6) with a highly conservated ankyrin repeat and a ’TRP domain’ [16][22]. TRPA1 is activated by irritants and pungent agents (e.g., acrolein, AITC) through a reversible covalent modification of specific cysteine residues in the N-terminal [49][71]. Being an ion channel protein, a pore is formed between the S5 and S6 domains. Molecular modeling and sequence alignment revealed individual residues within the S5 and S6 segments, which are responsible for the opening and activation of the channel [43][72]. Despite the homology, TRP channels respond to a wide range of stimuli and possess diverse gating mechanisms, ion selectivity, and functions. However, the structural similarities may provide a basis for interaction between distinct TRP receptors when co-expressed [50][78]. The cooperation between TRPA1 and TRPV1 is suggested by a high-order co-expression of the two TRP receptors. It was earlier described that nearly all TRPA1-sensitive neurons also express TRPV1 [17][23]. Functional studies proved that the two channels can co-work in a synergistic manner [43][44][72,73]. Interestingly, the inflammatory mediator, bradykinin, has been found to activate both TRPA1 and TRPV1, and it seems that both receptors are required for the sensory neurons’ maximum response elicited by bradykinin [45][74]. The TRPA1 channel activity has been shown to be regulated and modulated by the expression of TPRV1 in trigeminal neurons [51][79]. Novel research has revealed possible cooperation between TRPA1 and TRPV1 by attaching distinct subunits to create structurally atypical heteromers. Fisher et al. created TRPA1:TRPV1 complexes in cultured sensory neurons. The heteromers that formed were found to be less sensitive to TRPV1 agonists; therefore, it was concluded that TRPV1 activity is inhibited by its combination with TRPA1 subunits [52][80]. TRPA1 function also appears to be negatively regulated by the complex formation, as TRPA1:TRPV1 heteromers failed to respond to TRPA1 agonists. All TRPs are intended to form tetrameric assemblies, but different TRP subfamilies are able to combine their subunits to form heterotetramer complexes. Although the combination of TRP subunits is not so obvious, the interaction is provided by special modules in the transmembrane domains [53][81]. The research group also identified a unique domain on the intracellular terminals of TRPV1 that is thought to be responsible for the formation of the TRPA1:TRPV1 complex. The ablation of the identified residues reduced the possibility of the TRPA1:TRPV1 interaction. Tmem100, a conserved transmembrane protein, has been identified as a key regulator of TRPA1:TRPV1 cooperation [54][82]. This impressive study demonstrated that Tmem100 can physically interact with both TRPA1 and TRPV1 and reduce their association. Tmem100 appears to regulate TRPA1 activity in a TRPV1-dependent manner: in the absence of TRPV1, Tmem100 reduced the activity of the TRPA1. Weng et al. concluded that Tmem100 enhances TRPA1 activity by lowering the potential of TRPV1 to form complexes. They designed a peptide that blocks Tmem100 and consequently inhibits TRPA1 activity. The method could provide specific selective suppression of TRPV1- and TPRA1-mediated pain. The functional significance of the TRPA1:TRPV1 complex in the nociceptive pathway is under ongoing investigation and has not yet been clarified. Such physical interactions between two receptors can alter the receptor properties and excitability, so the cooperation of TRPA1 and TRPV1 should be carefully considered in chronic pain-related disease, such as in migraine, studies (Figure 1).