Coronavirus infections are neuroinvasive and can provoke injury to the central nervous system (CNS) and long-term illness consequences. They may be associated with inflammatory processes due to cellular oxidative stress and an imbalanced antioxidant system. The ability of phytochemicals with antioxidant and anti-inflammatory activities, such as Ginkgo biloba, to alleviate neurological complications and brain tissue damage has attracted strong ongoing interest in the neurotherapeutic management of long COVID. Ginkgo biloba leaf extract (EGb) contains several bioactive ingredients, e.g., bilobalide, quercetin, ginkgolides A–C, kaempferol, isorhamnetin, and luteolin. They have various pharmacological and medicinal effects, including memory and cognitive improvement. Ginkgo biloba, through its anti-apoptotic, antioxidant, and anti-inflammatory activities, impacts cognitive function and other illness conditions like those in long COVID.
- Ginkgo biloba bioactive compounds
- neuroinvasive coronavirus infection
- neurological long COVID
1. Introduction
2. Long-Term Neurological Damage and the Role of Oxidative Stress
2.1. Oxidative Stress and Redox Signaling, Players in SARS-CoV-2 Neurological Damage
It has been shown that Parkinson’s disease and SARS-CoV-2 infection cause similar oxidative stress and the activation of nuclear factor kappa B (NF-κB). In addition, activating pro-inflammatory mediators, such as IL-1 and IL-6, can lead to amyloid beta (Aβ) deposition and accumulation, thus establishing a link between neuroinflammation and Alzheimer’s disease [21][22][23][24]. It has been demonstrated that the receptor-binding domain (RBD) of the SARS-CoV-2 spike 1 (S1) protein binds to Aβ and tau proteins and causes their aggregation [25]. Additionally, SARS-CoV-2 infection has been linked to hypoxia and decreased oxygen levels [26]. The mitochondria of brain cells may undergo a rise in anaerobic metabolism due to this process, which can increase the levels of lactic acid, lipid peroxides, and oxygen-free radicals and deplete the antioxidant system. Consequently, the BBB is compromised, which may result in CNS complications. In regard to the role of oxidative stress in chronic diseases, Aranda-Rivera et al. highlighted the importance of nuclear factor erythroid 2–related factor 2 (Nrf2), a ubiquitous protein (containing 605 amino acids) that can modulate cellular oxidative stress [27]. During oxidative stress, Keap1 undergoes conformational change due to the oxidation of residues Cys 226, Cys 613, and Cys 624 by electrophiles and oxidants [28]. This mechanism enables Nrf2 to escape ubiquitination, leading to its release into the nucleus and regulating the expression of a number of antioxidant and detoxifying enzymes, such as glutamate-cysteine ligase modifier (GCLM) subunits, heme oxygenase, NAD(P)H quinone dehydrogenase 1 (NQO1), glutathione S-transferase, and glutathione peroxidase [29][30]. It should also be emphasized that viral infections, such as SARS-CoV-2, have adverse effects on antioxidant systems and have been linked to phenomena involving the inhibition of Nrf2 and activation of the NF-kB pathway in favor of inflammation and oxidative stress. Olagnier et al. showed that during SARS-CoV-2 infection, the Nrf2 pathway is repressed, leading to the downregulation of heme oxygenase 1 (HO-1) and NQO1 [31]. Therefore, several antioxidant enzymes that guard against oxidative stress, including glutathione peroxide, peroxiredoxin, thioredoxin reductase, and thioredoxin, are affected. Numerous studies have shown that the phosphatidylinositol 3-kinase (PI3K)/AKT signaling pathway controls oxidative stress by activating the transcription factor FOXO3 and initiating the transcription of antioxidant proteins, such as SOD-2, peroxiredoxins (PRDXs) 3 and 5, which are found in the mitochondria, and catalase, which is found in the peroxisomes [32][33][34]. Peroxisome-proliferator-activated receptor coactivator 1 (PGC-1), the master biogenesis regulator that promotes the transcription of antioxidant enzymes, interacts with the FOXO3 transcription factor to control oxidative stress in the mitochondria [33]. However, research has revealed that the ability of PGC-1 to promote gluconeogenesis and fatty acid oxidation is inhibited by protein kinase Akt2/protein kinase B (PKB), which acts as an intermediary trigger of phosphorylation and inhibition [35]. NF-κB is another transcription factor that regulates stress responses. It is activated by the phosphorylation of I-κB, thanks to the I-κB kinase (IKK) complex. In vitro studies by Wu et al. have reported that sustained exposure of human lens epithelial cells (HLECs) to increasing doses of H2O2 (50–100 μM) for 4 h attenuates the TNFα-induced degradation of I-κB, accompanied by the activation of NF-κB and proteasome activity by 50–80% [36]. The obtained data have also indicated that the activation of NF-κB is an essential phenomenon that enables cells to recover from oxidative stress.2.2. In Vivo and In Vitro Models of Oxidative Stress
Over the years, various oxidative stress models have been developed to study the pathogenesis of neurodegenerative disorders and discover new strategies for developing therapeutic agents. Such experimental techniques include the use of lipid peroxidation products, endogenous antioxidant depletion, mitochondrial respiratory chain inhibitors, neurotoxic agents (e.g., rotenone and N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)), and ischemic brain damage models [22]. The proposed in vitro cellular models and in vivo animal models have shed light on the molecular mechanisms underlying oxidative stress responses in the nervous system, such as cell survival and cell death. Among neurotoxic chemical agents, 6-hydroxydopamine (6-OHDA) has been used to induce neurotoxicity in the dopaminergic nigrostriatal system by inhibiting the mitochondrial electron transport chain of complexes I and IV and accelerating neuronal degeneration [37][38][39]. 6-OHDA has been regarded as an endogenous toxic factor in the pathogenesis of PD. The neurotoxin 6-OHDA induces excessive production and accumulation of ROS and, therefore, oxidative stress. In fact, 6-OHDA induces caspase-3 activation in the cells mediated by the Fas or mitochondrial pathways [40]. Indeed, it has been demonstrated that MTPT/6-OHDA-induced NF-κB activation in SH-SY5Y neuroblastoma cells triggers caspase-3 activation, which results in the death of DCNs via the NF-κB pathway [41][42][43]. In vitro studies have evaluated the toxic effects of 6-OHDA in dopaminergic (DArgic) cell cultures. For instance, Vestuto et al. used human neuroblastoma SH-SY5Y cells to assess the neuroprotective effect of cocoa extract (purified fractions) in a 6-OHDA-induced PD cellular model [44]. Similarly, Chansiw et al. reported the protective effect of 1-(N-acetyl-6-aminohexyl)-3-hydroxy-2-methyl pyridine-4-one (CM1) coupled with green tea extract (GTE) on iron-induced oxidative stress in SH-SY5Y cells [45]. In a separate study, Chen et al. evaluated the protective effect of EGCG against 6-OHDA-induced neurotoxicity by using N27 dopaminergic neurons [46]. The advantages of cellular systems for studying oxidative stress are their low cost, adaptability, modularity, reproducibility, compatibility with high-throughput screening, and interest in cell mechanical investigations without systemic interferences. Other studies have demonstrated that hypoxia can increase cells’ susceptibility to oxidative stress. The hypoxia-reoxygenation model is a relevant in vitro model of oxidative stress, given that cellular hypoxia appears to be a crucial signal that activates transcriptional regulators, specifically hypoxia-inducible factor-1 (HIF-1), nuclear factor kappa B (NF-κB), activator protein 1 (AP-1), and some mitogen-activated protein kinase (MAPK) signaling pathways, and induces cell death and necrosis [47][48][49][50]. Genetically derived models of neurodegenerative diseases are gaining considerable interest because they are excellent surrogates, providing intrinsic validity to genetically based models of degenerative disorders [51]. Scientific investigations have reported that knockout of genes, including PINK1, DJ-1, LRRK2, and LRRK1, in rats leads to age-dependent neurodegeneration of the dopaminergic neurons of PD [52][53][54]. Moreover, mutation in the copper-zinc superoxide dismutase 1 (SOD1) gene has been associated with ALS, while the alteration of MAPT genes or progranulin is linked to frontotemporal dementia (FTD) [55]. Other studies have highlighted that mutations in amyloid precursor protein (APP), presenilin 1, and presenilin 2 (PSEN1/2) are the main causes of autosomal dominant early-onset AD [56][57]. It has been documented that invertebrates can also mimic endogenous-generated ROS. Such a model has been widely implemented in Drosophila melanogaster and Caenorhabditis elegans (C. elegans) [58][59]. The initial longevity mutant, known as age-1, which contains increased levels of both SOD1 and catalase, is arguably the best-studied mutant in the nematode C. elegans. The age-1 mutant exhibits more significant levels of both SOD1 and catalase enzymes because this gene, which encodes for phosphatidylinositol-3 kinase, confers a longer lifetime phenotype when silenced, along with improved resilience to several forms of stress [59][60][61].3. Ginkgo Biloba Extract (EGb) for Neuroprotection and Potential Regeneration from Long COVID Syndrome
3.1. Ginkgo Biloba Antioxidative and Anti-Inflammatory Effects
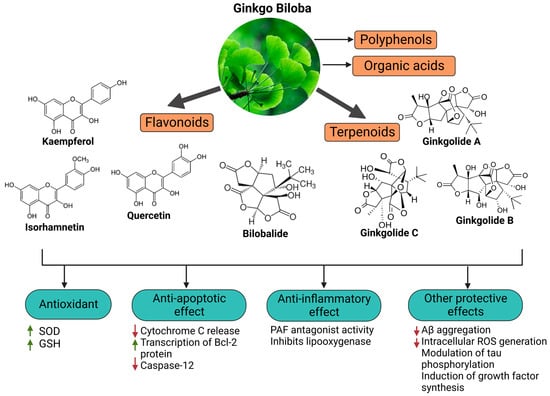
Ginkgo biloba (GB) is one of the medicinal plants that ameliorate capillary blood circulation, provide brain oxygenation, and thus improve age-related disorders. The ginkgo tree is monotypic and belongs to the class Ginkgoopsida, considered the oldest tree alive in the world (ginkgo species are from the Permian period, around 286–248 million years ago). The currently available herbal medicines based on Ginkgo biloba extract (EGb) are Tebonin® and Tanakan®, which are mostly standardized on ginkgo flavone glycosides and terpene lactones—EGb 761® [17][20]. The standardized extract of Ginkgo biloba leaves includes 6% terpenoids, of which 3.1% are ginkgolides A, B, C, and J and 2.9% are bilobalide. It contains 24% flavonoid glycosides, including quercetin, kaempferol, and isorhamnetin, and 5–10% organic acids (Figure 1) [62]. Ginkgo biloba leaf extract is mentioned in the British Herbal Compendium as a treatment for mild-to-severe dementia, including Alzheimer’s disease, and for the treatment of neurological symptoms attributed to loss of concentration and poor memory, confusion, depression, anxiety, vertigo, tinnitus, and headache [63]. The neuroprotective effect of EGb 761 extract has been examined in a rat model of cerebral injury following ischemia/reperfusion (I/R). The treatment resulted in a decrease in MDA levels, the downregulation of pro-inflammatory cytokines (TNF-α and IL-1β), and an increase in the expression of anti-inflammatory cytokines (IL-10) and enzymatic SOD and myeloperoxidase (MPO) activities, which can control neurological impairments. It can be suggested that the beneficial effects of EGb on cerebral ischemia/reperfusion I/R injury result from the reduction in oxidative stress due to the inhibition of nitric oxide production and inflammation induced by I/R [64]. Kaempferol is one of the most important constituents of Ginkgo biloba, and its action accounts for the upregulation of the glutamate-cysteine ligase catalytic (GCLC) subunit, brain-derived neurotrophic factor (BDNF), B-cell lymphoma protein 2 (Bcl-2), and GSH [65][66][67]. Kaempferol inhibits ROS generation by scavenging free radicals and efficiently protects neuronal cells from oxidative injury. Additionally, it inhibits the production of pro-apoptotic proteins, including Bax and caspase-3, and modulates the downregulation of the NF-κB pathway to exert anti-apoptotic effects . A study by Zhou et al. showed that kaempferol can inhibit mitochondrial membrane transition (mPTP) opening and suppresses the release of cytochrome C via GSK-3β inhibition. Kaempferol is also involved in the inhibition of serotonin breakdown by monoamine oxidase, reduces neurotoxicity induced by 3-nitropropionic acid (3-NP), and induces the upregulation of heme oxygenase 1 (HMOX-1) [68][69][70]. Quercetin, bilobalide, and isorhamnetin are other essential compounds in Ginkgo biloba extracts. Bilobalide decreases the expression of reactive species induced through H2O2, thus inhibiting ER stress . It can also suppress pro-inflammatory activation, NF-κB, and COX-2 activities [71]. Studies have reported the beneficial effects of bilobalide in the upregulation of c-myc and p53 proteins, inhibition of the degradation of membrane phospholipids, and increased cellular proliferation of neurons in the hippocampus [72][71][73][74]. According to Wang et al., pretreatment with bilobalide substantially reduced COX-2, iNOS, and phosphorylated p65 in sepsis-induced CLP mouse models, while inducing I-kB activation in the lungs. Additionally, bilobalide reduced oxidative stress by increasing HO-1 expression in lung tissues and antioxidative enzyme genes, including catalase, MnSOD, CuZnSOD, and GPx-1 [73]. Moreover, bilobalide and EGb50 can modulate the expression of TLR4, NF-B, and MyD88, preventing the onset of acute lung injury (ALI) [73][74][75][76]. Ginkgo biloba components may therefore prevent the onset of ALI and the cytokine storm syndrome in COVID-19 by inhibiting pro-inflammatory signaling via the NF-κB and TLR4 signaling pathways. A study showed that the administration of EGb50 substantially lowers TNF- and IL-1 levels and prevents the related signal transduction through the p38 MAPK and NF-B p65 pathways in LPS-stimulated microglial cells [77]. In a separate study, EGb50 demonstrated a potential anti-inflammatory action by suppressing NLRP3-inflammasome-induced microglial activation [78]. For comparison, isorhamnetin has been linked with the inhibition of apoptosis and the suppression of DNA fragmentation [79]. In an enzymatic inhibition assay, it was established that ginkgolide A can act as an irreversible inhibitor against SARS-CoV-2 papain-like protease (PLpro) at a nontoxic dose of 1.79 µM [80]. Similarly, quercetin, the primary EGb flavonoid component, inhibits SARS-CoV-2 3-chymotrypsin-like protease (3CLpro) and PLpro, with a corresponding docking energy of 6.25–4.62 kcal/mol, preventing SARS-CoV-2 replication [81]. In a recent study, Liu et al. compared the antioxidant capacity of various Ginkgo biloba extracts by evaluating the mechanisms of ginkgolides A (GA), B (GB), K (GK), and bilobalide (BB) against oxidative stress caused by transient focal cerebral ischemia [82]. In vivo studies have been performed in a developed middle cerebral artery occlusion (MCAO) model of cerebral ischemic injury using male SD rats, followed by reperfusion and Ginkgo biloba treatments [82]. Neuroblastoma cells (SH-SY5Y) were subjected to oxygen-glucose deprivation (OGD) for 4 h, followed by 6 h of reoxygenation using ginkgolides and bilobalide. The in vitro experimental findings revealed that GA, GB, GK, and BB significantly reduce ROS and increase SOD activities and protein levels, including HO-1 and Nqo1. Additionally, p-Akt and p-Nrf2 levels considerably increased following ginkgolide and BB treatments, with GB demonstrating greater efficacy than GA and GK. These upregulations could be reduced in a dose-dependent manner by LY294002, a PI3K inhibitor [82].3.2. Neuroprotective, Anti-Apoptotic, and Anxiolytic Drug Effects of EGb
Results from microarray experiments have established the neuroprotective effect of Egb 761 against ischemic-induced neuronal injury. The data revealed that the upregulation of Bcl-2 protein may be mediated by the activation of cAMP-response-element-binding protein (CREB) [83]. EGb 761 increases CREB phosphorylation via the activation of PI3K/Akt and extracellular-signal-regulated kinase (ERK) signaling pathways [84]. The consequently released BNDF protects the neurons against ischemia. Moreover, Tchantchou et al. affirmed that EGb 761 can reduce Aβ oligomerization and promote neurogenesis by the phosphorylation of CREB. This was evidenced by enhanced cell proliferation in the hippocampus of TgAPP/PS1 mice [85]. Overall, these studies have demonstrated that flavonoids, the primary active constituents of EGb 761, may upregulate the CREB–BDNF pathway and therefore exert neuroprotection. Combination therapy of EGb 761 with bone-marrow-derived mesenchymal stem cells (BMSCs) has shown a synergistic effect in animals with autoimmune encephalomyelitis. The therapeutic mechanism involves the inhibition of pro-inflammatory cytokines, demyelination, and protection axons and neurons [86]. Other studies have documented that GA prevents p-Tau deposition and thus protects cells from toxicity associated with Tau hyperphosphorylation. Interestingly, the degree of dementia is closely correlated with the production of hyperphosphorylated Tau aggregates, making EGb 761 crucial to counteract the neurodegenerative process [87]. Ginkgo biloba extracts, mainly flavonoids and ginkgolides, exert inhibitory effects on acetylcholinesterase activity. In fact, cholinergic agonists can reduce inflammation by blocking inflammatory signals, particularly the ubiquitous nuclear protein HMGB1 that is released by dying cells or activated innate immunity cells to promote inflammation [88]. It has been suggested that nicotinic receptors nAChRs may control the expression of ACE2 and serve as a binding receptor for S1 protein, leading to an inflammatory response. However, EGb may counteract the central inhibitory and anti-inflammatory effects of GABAergic neurons, leading to increased cortical neuronal activity and an increased risk of convulsion [89][90]. However, meta-analysis research has proven that there is no convulsion risk associated with the anxiolytic action of EGb, which is mediated through the regulation of GABAergic neurons. In patients with dementia, EGb 761® has shown promise in alleviating comorbid neurosensory symptoms and improving memory deficits [91]. In another study, EGb 761 showed neuroprotective effects against oxidative-stress-induced apoptotic cell death by inhibiting apoptosis in a p53-dependent pathway, preventing mitochondrial membrane damage, reducing the release of cytochrome C from the mitochondria, upregulating the anti-apoptotic protein Bcl-2, and inhibiting PARP cleavage [92]. The anti-apoptotic effects are schematically presented in Figure 2.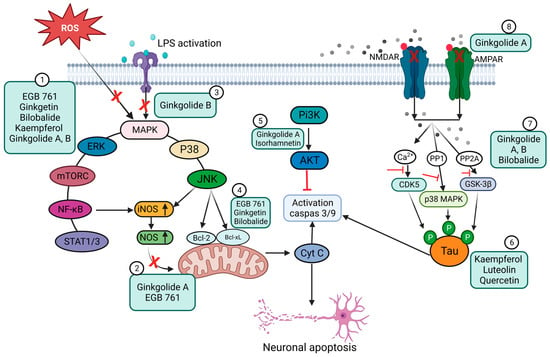
4. Ginkgo-Biloba-Based Nanotherapy for Neuroprotection and Regeneration from SARS-CoV-2 Neurological Damage
5. Conclusions
It is indispensable to continue studying the mechanisms that underlie the pathophysiological process of SARS-CoV-2 infection. This will enable researchers to uncover the therapeutic targets that may be used for their management. According to this resviearchw, it may be suggested that Ginkgo biloba has potential positive effects, including anxiolytic, antineurotoxic, anti-inflammatory and anti-apoptotic functions, and has been explored in treating neurological disorders, particularly AD, PD, and dementia. Nevertheless, further studies are needed to corroborate the activity and mechanisms of action of this phytochemical since it could constitute an alternative for the treatment of vascular and degenerative diseases. Nanotechnology-based drug delivery systems could be an approach to address the limitations of antioxidant compounds, which include insufficient dosing, limited bioavailability, restricted transport to the CNS, transient retention, and low antioxidant capacity to completely scavenge the effect of ROS. The development of experimental techniques to mimic ROS has made it possible to study oxidative stress in the CNS. These methods will be fundamental for future discoveries related to the role of oxidative stress in neurological diseasesReferences
- Raveendran, A.V.; Jayadevan, R.; Sashidharan, S. Long COVID: An Overview. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 869–875.
- Wieczfinska, J.; Kleniewska, P.; Pawliczak, R. Oxidative Stress-Related Mechanisms in SARS-CoV-2 Infections. Oxidative Med. Cell. Longev. 2022, 2022, 5589089.
- Kase, Y.; Okano, H. Neurological Pathogenesis of SARS-CoV-2 (COVID-19): From Virological Features to Clinical Symptoms. Inflamm. Regen. 2021, 41, 15.
- Nuzzo, D.; Picone, P. Potential Neurological Effects of Severe COVID-19 Infection. Neurosci. Res. 2020, 158, 1–5.
- Edinoff, A.N.; Chappidi, M.; Alpaugh, E.S.; Turbeville, B.C.; Falgoust, E.P.; Cornett, E.M.; Murnane, K.S.; Kaye, A.M.; Kaye, A.D. Neurological and Psychiatric Symptoms of COVID-19: A Narrative Review. Psychiatry Int. 2022, 3, 158–168.
- Sykes, D.L.; Holdsworth, L.; Jawad, N.; Gunasekera, P.; Morice, A.H.; Crooks, M.G. Post-COVID-19 Symptom Burden: What Is Long-COVID and How Should We Manage It? Lung 2021, 199, 113–119.
- Ahamed, J.; Laurence, J. Long COVID Endotheliopathy: Hypothesized Mechanisms and Potential Therapeutic Approaches. J. Clin. Investig. 2022, 132, e161167.
- Carfì, A.; Bernabei, R.; Landi, F.; for the Gemelli Against COVID-19 Post-Acute Care Study Group. Persistent Symptoms in Patients After Acute COVID-19. JAMA 2020, 324, 603.
- Pilotto, A.; Cristillo, V.; Cotti Piccinelli, S.; Zoppi, N.; Bonzi, G.; Sattin, D.; Schiavolin, S.; Raggi, A.; Canale, A.; Gipponi, S.; et al. Long-Term Neurological Manifestations of COVID-19: Prevalence and Predictive Factors. Neurol. Sci. 2021, 42, 4903–4907
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological Findings of COVID-19 Associated with Acute Respiratory Distress Syndrome. Lancet Respir. Med. 2020, 8, 420–422
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.-C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System: Celebrating the 20th Anniversary of the Discovery of ACE2. Circ. Res. 2020, 126, 1456–1474.
- Villa, C.; Rivellini, E.; Lavitrano, M.; Combi, R. Can SARS-CoV-2 Infection Exacerbate Alzheimer’s Disease? An Overview of Shared Risk Factors and Pathogenetic Mechanisms. JPM 2022, 12, 29.
- Domingues, R.B.; Mendes-Correa, M.C.; de Moura Leite, F.B.V.; Sabino, E.C.; Salarini, D.Z.; Claro, I.; Santos, D.W.; de Jesus, J.G.; Ferreira, N.E.; Romano, C.M.; et al. First Case of SARS-COV-2 Sequencing in Cerebrospinal Fluid of a Patient with Suspected Demyelinating Disease. J. Neurol. 2020, 267, 3154–3156
- Xiong, Y.; Zhu, G.-H.; Wang, H.-N.; Hu, Q.; Chen, L.-L.; Guan, X.-Q.; Li, H.-L.; Chen, H.-Z.; Tang, H.; Ge, G.-B. Discovery of Naturally Occurring Inhibitors against SARS-CoV-2 3CLpro from Ginkgo Biloba Leaves via Large-Scale Screening. Fitoterapia 2021, 152, 104909.
- Chen, Z.; Cui, Q.; Cooper, L.; Zhang, P.; Lee, H.; Chen, Z.; Wang, Y.; Liu, X.; Rong, L.; Du, R. Ginkgolic Acid and Anacardic Acid Are Specific Covalent Inhibitors of SARS-CoV-2 Cysteine Proteases. Cell Biosci. 2021, 11, 45.
- Sochocka, M.; Ochnik, M.; Sobczyński, M.; Gębura, K.; Zambrowicz, A.; Naporowski, P.; Leszek, J. Ginkgo Biloba Leaf Extract Improves an Innate Immune Response of Peripheral Blood Leukocytes of Alzheimer’s Disease Patients. Nutrients 2022, 14, 2022
- Barbalho, S.M.; Direito, R.; Laurindo, L.F.; Marton, L.T.; Guiguer, E.L.; Goulart, R.D.A.; Tofano, R.J.; Carvalho, A.C.A.; Flato, U.A.P.; Capelluppi Tofano, V.A.; et al. Ginkgo Biloba in the Aging Process: A Narrative Review. Antioxidants 2022, 11, 525.
- Tomino, C.; Ilari, S.; Solfrizzi, V.; Malafoglia, V.; Zilio, G.; Russo, P.; Proietti, S.; Marcolongo, F.; Scapagnini, G.; Muscoli, C.; et al. Mild Cognitive Impairment and Mild Dementia: The Role of Ginkgo Biloba (EGb 761®). Pharmaceuticals 2021, 14, 305.
- Singh, S.K.; Srivastav, S.; Castellani, R.J.; Plascencia-Villa, G.; Perry, G. Neuroprotective and Antioxidant Effect of Ginkgo Biloba Extract Against AD and Other Neurological Disorders. Neurotherapeutics 2019, 16, 666–674.
- Al-kuraishy, H.M.; Al-Gareeb, A.I.; Kaushik, A.; Kujawska, M.; Batiha, G.E. Ginkgo Biloba in the Management of the COVID-19 Severity. Arch. Pharm. 2022, 355, 2200188.
- Sulzer, D.; Antonini, A.; Leta, V.; Nordvig, A.; Smeyne, R.J.; Goldman, J.E.; Al-Dalahmah, O.; Zecca, L.; Sette, A.; Bubacco, L.; et al. COVID-19 and Possible Links with Parkinson’s Disease and Parkinsonism: From Bench to Bedside. NPJ Park. Dis. 2020, 6, 18.
- Chaudhry, Z.; Klenja, D.; Janjua, N.; Cami-Kobeci, G.; Ahmed, B. COVID-19 and Parkinson’s Disease: Shared Inflammatory Pathways Under Oxidative Stress. Brain Sci. 2020, 10, 807.
- Serrano-Castro, P.J.; Estivill-Torrús, G.; Cabezudo-García, P.; Reyes-Bueno, J.A.; Ciano Petersen, N.; Aguilar-Castillo, M.J.; Suárez-Pérez, J.; Jiménez-Hernández, M.D.; Moya-Molina, M.Á.; Oliver-Martos, B.; et al. Impact of SARS-CoV-2 Infection on Neurodegenerative and Neuropsychiatric Diseases: A Delayed Pandemic? Neurología 2020, 35, 245–251.
- Cavezzi, A.; Troiani, E.; Corrao, S. COVID-19: Hemoglobin, Iron, and Hypoxia beyond Inflammation. A Narrative Review. Clin. Pract. 2020, 10, 1271
- Idrees, D.; Kumar, V. SARS-CoV-2 Spike Protein Interactions with Amyloidogenic Proteins: Potential Clues to Neurodegeneration. Biochem. Biophys. Res. Commun. 2021, 554, 94–98.
- Cavezzi, A.; Troiani, E.; Corrao, S. COVID-19: Hemoglobin, Iron, and Hypoxia beyond Inflammation. A Narrative Review. Clin. Pract. 2020, 10, 1271.
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Pedraza-Chaverri, J.; Scholze, A. Nrf2 Activation in Chronic Kidney Disease: Promises and Pitfalls. Antioxidants 2022, 11, 1112.
- Horie, Y.; Suzuki, T.; Inoue, J.; Iso, T.; Wells, G.; Moore, T.W.; Mizushima, T.; Dinkova-Kostova, A.T.; Kasai, T.; Kamei, T.; et al. Molecular Basis for the Disruption of Keap1–Nrf2 Interaction via Hinge & Latch Mechanism. Commun. Biol. 2021, 4, 576.
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Arancibia-Hernández, Y.L.; Hernández-Cruz, E.Y.; Pedraza-Chaverri, J. RONS and Oxidative Stress: An Overview of Basic Concepts. Oxygen 2022, 2, 437–478.
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell. Biol. 2020, 40, e00099-20.
- Olagnier, D.; Farahani, E.; Thyrsted, J.; Blay-Cadanet, J.; Herengt, A.; Idorn, M.; Hait, A.; Hernaez, B.; Knudsen, A.; Iversen, M.B.; et al. SARS-CoV2-Mediated Suppression of NRF2-Signaling Reveals Potent Antiviral and Anti-Inflammatory Activity of 4-Octyl-Itaconate and Dimethyl Fumarate. Nat. Commun. 2020, 11, 4938.
- Klotz, L.-O.; Sánchez-Ramos, C.; Prieto-Arroyo, I.; Urbánek, P.; Steinbrenner, H.; Monsalve, M. Redox Regulation of FoxO Transcription Factors. Redox Biol. 2015, 6, 51–72.
- Borniquel, S.; García-Quintáns, N.; Valle, I.; Olmos, Y.; Wild, B.; Martínez-Granero, F.; Soria, E.; Lamas, S.; Monsalve, M. Inactivation of Foxo3a and Subsequent Downregulation of PGC-1α Mediate Nitric Oxide-Induced Endothelial Cell Migration. Mol. Cell. Biol. 2010, 30, 4035–4044.
- Li, X.; Monks, B.; Ge, Q.; Birnbaum, M.J. Akt/PKB Regulates Hepatic Metabolism by Directly Inhibiting PGC-1α Transcription Coactivator. Nature 2007, 447, 1012–1016.
- Andrzejewski, K.; Jampolska, M.; Zaremba, M.; Joniec-Maciejak, I.; Boguszewski, P.M.; Kaczyńska, K. Respiratory Pattern and Phrenic and Hypoglossal Nerve Activity during Normoxia and Hypoxia in 6-OHDA-Induced Bilateral Model of Parkinson’s Disease. J. Physiol. Sci. 2020, 70, 16
- Wu, M.; Bian, Q.; Liu, Y.; Fernandes, A.; Taylor, A.; Pereira, P.; Shang, F. Sustained Oxidative Stress Inhibits NF-ΚB Activation Partially via Inactivating the Proteasome. Free Radic. Biol. Med. 2009, 46, 62–69
- Andrzejewski, K.; Jampolska, M.; Zaremba, M.; Joniec-Maciejak, I.; Boguszewski, P.M.; Kaczyńska, K. Respiratory Pattern and Phrenic and Hypoglossal Nerve Activity during Normoxia and Hypoxia in 6-OHDA-Induced Bilateral Model of Parkinson’s Disease. J. Physiol. Sci. 2020, 70, 16. [Google Scholar]
- Deumens, R.; Blokland, A.; Prickaerts, J. Modeling Parkinson’s Disease in Rats: An Evaluation of 6-OHDA Lesions of the Nigrostriatal Pathway. Exp. Neurol. 2002, 175, 303–317.
- Glinka, Y.Y.; Youdim, M.B.H. Inhibition of Mitochondrial Complexes I and IV by 6-Hydroxydopamine. Eur. J. Pharmacol. Environ. Toxicol. Pharmacol. 1995, 292, 329–332
- Tanaka, K.; Ogawa, N.; Asanuma, M. Molecular Basis of 6-Hydroxydopamine-Induced Caspase Activations Due to Increases in Oxidative Stress in the Mouse Striatum. Neurosci. Lett. 2006, 410, 85–89
- Tirmenstein, M.A.; Hu, C.X.; Scicchitano, M.S.; Narayanan, P.K.; McFarland, D.C.; Thomas, H.C.; Schwartz, L.W. Effects of 6-Hydroxydopamine on Mitochondrial Function and Glutathione Status in SH-SY5Y Human Neuroblastoma Cells. Toxicol. Vitr. 2005, 19, 471–479.
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.-F.; Benabid, A.-L.; Sadoul, R.; Verna, J.-M. Molecular Pathways Involved in the Neurotoxicity of 6-OHDA, Dopamine and MPTP: Contribution to the Apoptotic Theory in Parkinson’s Disease. Prog. Neurobiol. 2001, 65, 135–172.
- Gong, P.; Deng, F.; Zhang, W.; Ji, J.; Liu, J.; Sun, Y.; Hu, J. Tectorigenin Attenuates the MPP+-induced SH-SY5Y Cell Damage, Indicating a Potential Beneficial Role in Parkinson’s Disease by Oxidative Stress Inhibition. Exp. Ther. Med. 2017, 14, 4431–4437.
- Vestuto, V.; Amodio, G.; Pepe, G.; Basilicata, M.G.; Belvedere, R.; Napolitano, E.; Guarnieri, D.; Pagliara, V.; Paladino, S.; Rodriquez, M.; et al. Cocoa Extract Provides Protection against 6-OHDA Toxicity in SH-SY5Y Dopaminergic Neurons by Targeting PERK. Biomedicines 2022, 10, 2009
- Chansiw, N.; Kulprachakarn, K.; Paradee, N.; Prommaban, A.; Srichairatanakool, S. Protection of Iron-Induced Oxidative Damage in Neuroblastoma (SH-SY5Y) Cells by Combination of 1-(N-Acetyl-6-Aminohexyl)-3-Hydroxy-2-Methylpyridin-4-One and Green Tea Extract. Bioinorg. Chem. Appl. 2021, 2021, 5539666
- Chen, D.; Kanthasamy, A.G.; Reddy, M.B. EGCG Protects against 6-OHDA-Induced Neurotoxicity in a Cell Culture Model. Park. Dis. 2015, 2015, 843906
- Semenza, G.L. HIF-1: Mediator of Physiological and Pathophysiological Responses to Hypoxia. J. Appl. Physiol. 2000, 88, 1474–1480.
- Chazelas, P.; Steichen, C.; Favreau, F.; Trouillas, P.; Hannaert, P.; Thuillier, R.; Giraud, S.; Hauet, T.; Guillard, J. Oxidative Stress Evaluation in Ischemia Reperfusion Models: Characteristics, Limits and Perspectives. IJMS 2021, 22, 2366.
- Irani, K. Oxidant Signaling in Vascular Cell Growth, Death, and Survival: A Review of the Roles of Reactive Oxygen Species in Smooth Muscle and Endothelial Cell Mitogenic and Apoptotic Signaling. Circ. Res. 2000, 87, 179–183.
- Saikumar, P.; Dong, Z.; Weinberg, J.M.; Venkatachalam, M.A. Mechanisms of Cell Death in Hypoxia/Reoxygenation Injury. Oncogene 1998, 17, 3341–3349
- Klein, C.; Westenberger, A. Genetics of Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a008888
- Klein, C.; Westenberger, A. Genetics of Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a008888.
- Martin, I.; Dawson, V.L.; Dawson, T.M. Recent Advances in the Genetics of Parkinson’s Disease. Annu. Rev. Genom. Hum. Genet. 2011, 12, 301–325.
- Scott, L.; Dawson, V.L.; Dawson, T.M. Trumping Neurodegeneration: Targeting Common Pathways Regulated by Autosomal Recessive Parkinson’s Disease Genes. Exp. Neurol. 2017, 298, 191–201
- Ittner, L.M.; Halliday, G.M.; Kril, J.J.; Götz, J.; Hodges, J.R.; Kiernan, M.C. FTD and ALS—Translating Mouse Studies into Clinical Trials. Nat. Rev. Neurol. 2015, 11, 360–366
- Haass, C.; Strooper, B.D. The Presenilins in Alzheimer’s Disease--Proteolysis Holds the Key. Science 1999, 286, 916–919.
- Dawson, T.M.; Golde, T.E.; Lagier-Tourenne, C. Animal Models of Neurodegenerative Diseases. Nat. Neurosci. 2018, 21, 1370–1379.
- Johnson, T.E.; de Castro, E.; Hegi de Castro, S.; Cypser, J.; Henderson, S.; Tedesco, P. Relationship between Increased Longevity and Stress Resistance as Assessed through Gerontogene Mutations in Caenorhabditis Elegans. Exp. Gerontol. 2001, 36, 1609–1617.
- Melov, S. Animal Models of Oxidative Stress, Aging, and Therapeutic Antioxidant Interventions. Int. J. Biochem. Cell Biol. 2002, 34, 1395–1400.
- Larsen, P.L. Aging and Resistance to Oxidative Damage in Caenorhabditis Elegans. Proc. Natl. Acad. Sci. USA 1993, 90, 8905–8909.
- Johnson, T.E. Increased Life-Span of Age-1 Mutants in Caenorhabditis Elegans and Lower Gompertz Rate of Aging. Science 1990, 249, 908–912
- Bradley, P.R. (Ed.) British Herbal Compendium: A Handbook of Scientific Information on Widely Used Plant Drugs; British Herbal Medicine Association: Bournemouth, UK, 1992.
- Bradley, P.R. (Ed.) British Herbal Compendium: A Handbook of Scientific Information on Widely Used Plant Drugs; British Herbal Medicine Association: Bournemouth, UK, 1992
- Yang, Y.; Liu, P.; Chen, L.; Liu, Z.; Zhang, H.; Wang, J.; Sun, X.; Zhong, W.; Wang, N.; Tian, K.; et al. Therapeutic Effect of Ginkgo Biloba Polysaccharide in Rats with Focal Cerebral Ischemia/Reperfusion (I/R) Injury. Carbohydr. Polym. 2013, 98, 1383–1388
- Achete de Souza, G.; de Marqui, S.V.; Matias, J.N.; Guiguer, E.L.; Barbalho, S.M. Effects of Ginkgo Biloba on Diseases Related to Oxidative Stress. Planta Med. 2020, 86, 376–386.
- Trebatická, J.; Ďuračková, Z. Psychiatric Disorders and Polyphenols: Can They Be Helpful in Therapy? Oxidative Med. Cell. Longev. 2015, 2015, 248529.
- Malik, J.; Choudhary, S.; Kumar, P. Plants and Phytochemicals for Huntington′s Disease. Pharmacogn. Rev. 2013, 7, 81.
- Ren, J.; Lu, Y.; Qian, Y.; Chen, B.; Wu, T.; Ji, G. Recent Progress Regarding Kaempferol for the Treatment of Various Diseases (Review). Exp. Ther. Med. 2019, 18, 2759–2776.
- Zhou, M.; Ren, H.; Han, J.; Wang, W.; Zheng, Q.; Wang, D. Protective Effects of Kaempferol against Myocardial Ischemia/Reperfusion Injury in Isolated Rat Heart via Antioxidant Activity and Inhibition of Glycogen Synthase Kinase-3 β. Oxidative Med. Cell. Longev. 2015, 2015, 481405.
- Yang, C.-C.; Hsiao, L.-D.; Wang, C.-Y.; Lin, W.-N.; Shih, Y.-F.; Chen, Y.-W.; Cho, R.-L.; Tseng, H.-C.; Yang, C.-M. HO-1 Upregulation by Kaempferol via ROS-Dependent Nrf2-ARE Cascade Attenuates Lipopolysaccharide-Mediated Intercellular Cell Adhesion Molecule-1 Expression in Human Pulmonary Alveolar Epithelial Cells. Antioxidants 2022, 11, 782.
- Ma, T.; Lv, L.; Yu, Y.; Jia, L.; Song, X.; Xu, X.; Li, T.; Sheng, X.; Wang, H.; Zhang, J.; et al. Bilobalide Exerts Anti-Inflammatory Effects on Chondrocytes Through the AMPK/SIRT1/MTOR Pathway to Attenuate ACLT-Induced Post-Traumatic Osteoarthritis in Rats. Front. Pharmacol. 2022, 13, 783506.
- Zhou, L.J.; Zhu, X.Z. Reactive Oxygen Species-Induced Apoptosis in PC12 Cells and Protective Effect of Bilobalide. J. Pharmacol. Exp. Ther. 2000, 293, 982–988.
- Wang, F.; Huang, J.; Li, J.; Chen, K.; Zhang, X.; Zhang, Y.; Zhu, Y. Bilobalide, a Bioactive Compound on Sepsis-Induced Acute Lung Injury through Its Anti-Inflammatory and Antioxidative Activity. Pharmacogn. Mag. 2021, 17, 163.
- Tchantchou, F.; Lacor, P.N.; Cao, Z.; Lao, L.; Hou, Y.; Cui, C.; Klein, W.L.; Luo, Y. Stimulation of Neurogenesis and Synaptogenesis by Bilobalide and Quercetin via Common Final Pathway in Hippocampal Neurons. JAD 2009, 18, 787–798.
- Zhou, J.-M.; Gu, S.-S.; Mei, W.H.; Zhou, J.; Wang, Z.Z.; Xiao, W. Ginkgolides and Bilobalide Protect BV2 Microglia Cells against OGD/Reoxygenation Injury by Inhibiting TLR2/4 Signaling Pathways. Cell Stress Chaperones 2016, 21, 1037–1053.
- Jiang, H.; Qu, P. Effects of Ginkgo Biloba Leaf Extract on Local Renin-Angiotensin System through TLR4/NF-ΚB Pathway in Cardiac Myocyte. Exp. Ther. Med. 2017, 14, 5857–5862.
- He, G.; Yuan, C.; Hao, L.; Xu, Y.; Zhang, Z. GBE50 Attenuates Inflammatory Response by Inhibiting the P38 MAPK and NF-κB Pathways in LPS-Stimulated Microglial Cells. Evid. Based Complement. Altern. Med. 2014, 2014, 368598.
- Liu, F.-Q.; Gao, Q.; Wang, D.-D.; Zhang, Z.-X. Effects of GBE50 on LPS/ATP induced NLRP3 inflammasome activation in primary rat microglia. China J. Chin. Mater. Med. 2018, 43, 3346–3352
- Gong, G.; Guan, Y.-Y.; Zhang, Z.-L.; Rahman, K.; Wang, S.-J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A Review of Pharmacological Effects. Biomed. Pharmacother. 2020, 128, 110301
- Chen, Z.; Cui, Q.; Cooper, L.; Zhang, P.; Lee, H.; Chen, Z.; Wang, Y.; Liu, X.; Rong, L.; Du, R. Ginkgolic Acid and Anacardic Acid Are Specific Covalent Inhibitors of SARS-CoV-2 Cysteine Proteases. Cell Biosci. 2021, 11, 45
- Derosa, G.; Maffioli, P.; D’Angelo, A.; Di Pierro, F. A Role for Quercetin in Coronavirus Disease 2019 (COVID-19). Phytother. Res. 2021, 35, 1230–1236.
- Liu, Q.; Jin, Z.; Xu, Z.; Yang, H.; Li, L.; Li, G.; Li, F.; Gu, S.; Zong, S.; Zhou, J.; et al. Antioxidant Effects of Ginkgolides and Bilobalide against Cerebral Ischemia Injury by Activating the Akt/Nrf2 Pathway in Vitro and in Vivo. Cell Stress Chaperones 2019, 24, 441–452
- Smith, J.V.; Burdick, A.J.; Golik, P.; Khan, I.; Wallace, D.; Luo, Y. Anti-Apoptotic Properties of Ginkgo Biloba Extract EGb 761 in Differentiated PC12 Cells. Cell. Mol. Biol. 2002, 48, 699–707.
- Perianayagam, M.C.; Madias, N.E.; Pereira, B.J.G.; Jaber, B.L. CREB Transcription Factor Modulates Bcl2 Transcription in Response to C5a in HL-60-Derived Neutrophils. Eur. J. Clin. Investig. 2006, 36, 353–361
- Tchantchou, F.; Xu, Y.; Wu, Y.; Christen, Y.; Luo, Y. EGb 761 Enhances Adult Hippocampal Neurogenesis and Phosphorylation of CREB in Transgenic Mouse Model of Alzheimer’s Disease. FASEB J. 2007, 21, 2400–2408
- Hao, F.; Li, A.; Yu, H.; Liu, M.; Wang, Y.; Liu, J.; Liang, Z. Enhanced Neuroprotective Effects of Combination Therapy with Bone Marrow-Derived Mesenchymal Stem Cells and Ginkgo Biloba Extract (EGb761) in a Rat Model of Experimental Autoimmune Encephalomyelitis. Neuroimmunomodulation 2016, 23, 41–57
- Du, X.; Wang, X.; Geng, M. Alzheimer’s Disease Hypothesis and Related Therapies. Transl. Neurodegener. 2018, 7, 2.
- Andersson, U. The Cholinergic Anti-Inflammatory Pathway Alleviates Acute Lung Injury. Mol. Med. 2020, 26, 64.
- Ivic, L.; Sands, T.T.J.; Fishkin, N.; Nakanishi, K.; Kriegstein, A.R.; Strømgaard, K. Terpene Trilactones from Ginkgo Biloba Are Antagonists of Cortical Glycine and GABAA Receptors. J. Biol. Chem. 2003, 278, 49279–49285.
- Alexandris, N.; Lagoumintzis, G.; Chasapis, C.T.; Leonidas, D.D.; Papadopoulos, G.E.; Tzartos, S.J.; Tsatsakis, A.; Eliopoulos, E.; Poulas, K.; Farsalinos, K. Nicotinic Cholinergic System and COVID-19: In Silico Evaluation of Nicotinic Acetylcholine Receptor Agonists as Potential Therapeutic Interventions. Toxicol. Rep. 2021, 8, 73–83.
- Spiegel, R.; Kalla, R.; Mantokoudis, G.; Maire, R.; Mueller, H.; Hoerr, R.; Ihl, R. Ginkgo Biloba Extract EGb 761® Alleviates Neurosensory Symptoms in Patients with Dementia: A Meta-Analysis of Treatment Effects on Tinnitus and Dizziness in Randomized, Placebo-Controlled Trials. CIA 2018, 13, 1121–1127.
- Di Meo, F.; Cuciniello, R.; Margarucci, S.; Bergamo, P.; Petillo, O.; Peluso, G.; Filosa, S.; Crispi, S. Ginkgo Biloba Prevents Oxidative Stress-Induced Apoptosis Blocking P53 Activation in Neuroblastoma Cells. Antioxidants 2020, 9, 279
- Wang, L.P.; Zhang, X.Y.; Liu, N.; Ma, Z.Z.; Fang, D.S. Comparison of Integrated Traditional Chinese and Western Medicine Therapy on Vascular Cognitive Impairment with No Dementia. Genet. Mol. Res. 2015, 14, 4896–4902.
- Gschwind, Y.J.; Bridenbaugh, S.A.; Reinhard, S.; Granacher, U.; Monsch, A.U.; Kressig, R.W. Ginkgo Biloba Special Extract LI 1370 Improves Dual-Task Walking in Patients with MCI: A Randomised, Double-Blind, Placebo-Controlled Exploratory Study. Aging Clin. Exp. Res. 2017, 29, 609–619.
- Kuo, L.-C.; Song, Y.-Q.; Yao, C.-A.; Cheng, I.H.; Chien, C.-T.; Lee, G.-C.; Yang, W.-C.; Lin, Y. Ginkgolide A Prevents the Amyloid-β-Induced Depolarization of Cortical Neurons. J. Agric. Food Chem. 2019, 67, 81–89
- Karavelioglu, Z.; Cakir-Koc, R. Preparation of Chitosan Nanoparticles as Ginkgo Biloba Extract Carrier: In Vitro Neuroprotective Effect on Oxidative Stress-Induced Human Neuroblastoma Cells (SH-SY5Y). Int. J. Biol. Macromol. 2021, 192, 675–683
- Wang, L.; Zhao, X.; Yang, F.; Wu, W.; Liu, Y.; Wang, L.; Wang, L.; Wang, Z. Enhanced Bioaccessibility in Vitro and Bioavailability of Ginkgo Biloba Extract Nanoparticles Prepared by Liquid Anti-solvent Precipitation. Int. J. Food Sci. Technol
- Zhao, Y.; Xiong, S.; Liu, P.; Liu, W.; Wang, Q.; Liu, Y.; Tan, H.; Chen, X.; Shi, X.; Wang, Q.; et al. Polymeric Nanoparticles-Based Brain Delivery with Improved Therapeutic Efficacy of Ginkgolide B in Parkinson’s Disease. IJN 2020, 15, 10453–10467
- Dhawan, S.; Kapil, R.; Singh, B. Formulation Development and Systematic Optimization of Solid Lipid Nanoparticles of Quercetin for Improved Brain Delivery. J. Pharm. Pharmacol. 2011, 63, 342–351
- Haghighi, P.; Ghaffari, S.; Arbabi Bidgoli, S.; Qomi, M.; Haghighat, S. Preparation, Characterization and Evaluation of Ginkgo Biloba Solid Lipid Nanoparticles. Nanomed. Res. J. 2018, 3, 71–78
- Xu, Z.; Feng, Q.; Wang, M.; Zhao, H.; Lin, Y.; Zhou, S. Green Biosynthesized Silver Nanoparticles With Aqueous Extracts of Ginkgo Biloba Induce Apoptosis via Mitochondrial Pathway in Cervical Cancer Cells. Front. Oncol.
- Wang, Q.; Ma, R.; Liu, P.; Cheng, G.; Yang, Q.; Chen, X.; Wu, Z.; Yuan, D.; Chen, T. Efficient Sustained-Release Nanoparticle Delivery System Protects Nigral Neurons in a Toxin Model of Parkinson’s Disease. Pharmaceutics 2022, 14, 1731
- Xu, Z.; Feng, Q.; Wang, M.; Zhao, H.; Lin, Y.; Zhou, S. Green Biosynthesized Silver Nanoparticles With Aqueous Extracts of Ginkgo Biloba Induce Apoptosis via Mitochondrial Pathway in Cervical Cancer Cells. Front. Oncol.
- Wang, Q.; Ma, R.; Liu, P.; Cheng, G.; Yang, Q.; Chen, X.; Wu, Z.; Yuan, D.; Chen, T. Efficient Sustained-Release Nanoparticle Delivery System Protects Nigral Neurons in a Toxin Model of Parkinson’s Disease. Pharmaceutics 2022, 14, 1731