Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Young-Min Lee and Version 2 by Camila Xu.
Japanese encephalitis virus (JEV), a zoonotic flavivirus, is principally transmitted by hematophagous mosquitoes, continually between susceptible animals and incidentally from those animals to humans.
- Japanese encephalitis virus
- flavivirus
- pathogenesis
1. Introduction
Japanese encephalitis virus (JEV) is a mosquito-borne arbovirus taxonomically belonging to the genus Flavivirus in the family Flaviviridae [1]. Of the 53 classified species of known flaviviruses, JEV is related to other medically important mosquito-borne flaviviruses to various genetic and antigenic extents, with a much closer relationship being noted with West Nile (WNV), St. Louis encephalitis (SLEV), and Murray Valley encephalitis viruses than with Zika (ZIKV), dengue (DENV), and yellow fever (YFV) viruses [2][3][2,3]. JEV circulates in nature by both horizontal transmission among susceptible animal hosts, primarily through the bite of culicine mosquito vectors (e.g., Culex tritaeniorhynchus) [4][5][4,5], and vertical transmission from female mosquitoes to their offspring through the transovarial infection of developing eggs [6][7][6,7]. Of many susceptible animals, certain vertebrates, such as suids (e.g., domestic pigs) and avians (e.g., wading birds), are especially relevant for the incidental transmission of JEV to the human population [8][9][10][8,9,10]. Experimentally, JEV can also be transmitted in the absence of mosquito vectors through several non-vector-borne routes in animals such as rodents, pigs, bats, and/or squirrel monkeys: contact transmission [11][12][13][14][11,12,13,14], aerosol transmission [11][15][11,15], transplacental transmission [16][17][18][19][20][16,17,18,19,20], and artificial insemination [21][22][21,22]. In both humans and animals, JEV causes Japanese encephalitis (JE), formerly called Type B or Japanese B encephalitis [5][23][5,23], which is an acute encephalitis syndrome that potentially leads to severe brain damage or death [24][25][24,25]. In Japan, where JE was first recognized, the disease is believed to have arisen in the “summer encephalitis” season before the 20th century began, although the first recorded outbreak occurred in 1924, involving >6000 human cases with a case fatality of ~60% [26][27][26,27]. About a decade after this historic outbreak, a filterable agent from the brain of a fatal human JE case was demonstrated to be able to produce the disease in monkeys. In 1935, JEV was first isolated from the brain of a deceased patient, and the isolate was designated as the Nakayama strain [28]. Despite the fact that JEV has been known as the cause of JE for nearly a century, it is still a neglected pathogen that continues to be a major public health challenge [29].
JEV is the most common cause of viral encephalitis in the Asia-Pacific region [30][31][32][33][30,31,32,33], with the reported boundaries of viral activity on the north being much of China and the eastern China-Russia borderlands [34][35][36][37][38][39][40][34,35,36,37,38,39,40], on the south being Papua New Guinea [41][42][41,42] and a northern part of Australia [43][44][45][46][47][48][49][43,44,45,46,47,48,49], on the east being Guam [50][51][52][50,51,52] and Saipan [53][54][53,54], and on the west being much of India [55][56][55,56] and a southeastern part of Pakistan [57][58][57,58]. This region contains ~25 countries, including the top two most populous countries (i.e., China and India) of the world and several of the most densely populated countries (e.g., Singapore and Bangladesh), putting far more than half of the global population at risk of JEV infection [59]. Additionally, some cases of JEV infection were found in birds [60][61][60,61] and mosquitoes [62] collected in Italy during the first decade of the 21st century, which was the first time that the virus was detected outside the Asia-Pacific region, albeit with no human JE outbreaks reported as of yet. In 2016, however, a case of co-infection with JEV and YFV was identified unexpectedly in an Angolan resident with no out-of-country travel history [63]. This recent epidemiological data indicates that JEV is no longer confined to the Asia-Pacific region, and is continuously expanding its activity over new territories in Europe and Africa, with possible spread to the Americas in the very near future [64][65][64,65]. Hence, JEV is an emerging pathogen that has the potential to spread across the globe, now more than ever [66][67][68][66,67,68].
The annual incidence of JE presents two main epidemiological patterns [69][70][69,70], intrinsically aligned with the regional climate that affects the population size, density, and distribution of both mosquito vectors and animal hosts involved in JEV transmission [71][72][71,72]. In tropical regions, JEV is “endemic,” causing sporadic outbreaks nearly all year round at low frequencies, with peaks in the rainy season affecting all young under age 15 due to lack of immunity to the virus. In subtropical and temperate regions, on the other hand, JEV is “epidemic”, causing seasonal outbreaks exclusively during the summer and early fall and affecting all ages, with a bimodal distribution peaking in young children and the elderly because of the lack and the fading of JEV-specific immunity, respectively. Since its emergence, JEV has evolved into five divergent genotypes (G1–G5) presumably at the center of the Asia-Pacific region (i.e., Indonesia and Malaysia), from which one or more genotypes have dispersed to other areas of the region [73]. Historically, G1, G2, and G3 have all been circulated broadly at various times and frequencies in the Asia-Pacific region, except the India-Nepal-Sri Lanka area in which only G1 and G3 have been detected, and the Australia-Papua New Guinea area in which only G1 and G2 have been found [74][75][76][77][78][79][80][81][82][83][74,75,76,77,78,79,80,81,82,83]. Of particular note, G3 was the most prevalent in Asia until the 1990s, but since then, it has slowly been replaced by G1 [74][75][76][77][78][79][80][74,75,76,77,78,79,80]; yet, G3 has recently appeared for the first time outside the Asia-Pacific region in Italy [60][61][62][60,61,62] and Angola [63]. Unlike the three aforementioned genotypes, G4 has been located only in the Indonesia-Malaysia area [84], and G5 has been limited to the China-Japan-Korea area apart from the Indonesia-Malaysia area [85][86][87][88][85,86,87,88]. Within the five genotypes, it is noted that the sequence divergence between their genomes at the nucleotide (nt) and amino acid (aa) levels reaches up to ~20% and ~10%, respectively [85]; however, their biological differences in viral transmissibility, pathogenicity, and immunogenicity have not yet been fully understood.
2. The Virus
JEV is an enveloped virus with a linear, single-stranded, positive-sense RNA genome [89][100]. The virion consists of an inner nucleocapsid composed of the genomic RNA and multiple capsid C:C homodimers [90][101], and an outer shell composed of two surface proteins M and E arranged into 90 E:M:M:E heterotetramers [91][102] in a herringbone pattern on the viral envelope membrane (Figure 1, top panel). The genomic RNA is ~10,977 nt long (Figure 1, bottom panel) and has three functional parts [92][103]: (1) an ~95-nt untranslated region (UTR) with a type 1 cap structure attached at the 5’ end [93][104], (2) an ~10,299-nt central open reading frame (ORF) with a -1 ribosomal frameshift signal located at positions nt 3551-3630 [94][105], and (3) an ~583-nt UTR with no poly(A) tail at the 3’ end [95][106]. Each of the 5’ and 3’ UTRs, as well as the ~100-nt 5’-terminal region of the ORF, contains a network of cis-acting RNA elements [96][97][98][107,108,109] defined by the primary, secondary, and tertiary structures that regulate the translation and replication of the viral genomic RNA [99][100][101][102][110,111,112,113]. The single long ORF encodes two precursor polyproteins [103][114]: (i) the full-length ppC–NS5, which undergoes co- and post-translational proteolytic cleavages to yield three structural (C, prM/M, and E) and seven nonstructural (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) proteins; and (ii) the frameshift-derived truncated ppC–NS1’, which also goes through the same proteolysis as does ppC–NS5 to produce the three aforementioned structural proteins and NS1’, a 52-aa longer isoform of NS1 as a result of its C-terminal extension that includes the 9-aa N-terminal segment of NS2A before the frameshift and a 43-aa unique peptide synthesized from the new reading frame afterwards [92][104][105][103,115,116]. Of these 10 mature proteins, the following two are enzymes, each containing multiple catalytic activities [106][107][108][109][117,118,119,120]: (a) NS3, 619 aa in length, has serine protease activity in the ~168-aa N-terminal region [110][121] and helicase, nucleoside 5’-triphosphatase, and RNA 5’-triphosphatase activities in the ~438-aa C-terminal region [111][122], and (b) NS5, 905 aa in length, has methyltransferase and guanylyltransferase activities in the ~265-aa N-terminal region and RNA-dependent RNA polymerase activity in the ~630-aa C-terminal region [112][123]. In a nutshell, the three structural proteins are required for the formation of an infectious virion [113][114][115][124,125,126], and the seven nonstructural proteins are engaged in multiple steps of viral replication (i.e., polyprotein processing [116][127], RNA replication [117][118][119][128,129,130], and particle morphogenesis [120][121][131,132]) and in a variety of host cell responses to viral replication [122][123][124][125][133,134,135,136]. With regard to JEV pathogenesis, the viral E protein has been the target of extensive study since it acts as the viral receptor interacting with an ill-defined cellular component(s) on susceptible host cells for viral entry [126][127][128][137,138,139]. In addition, a recent study has suggested that a secreted form of the viral NS1 protein specifically binds to brain endothelial cells and alters their permeability, causing brain-specific vascular leakage that contributes to the pathogenesis of JE [129][140].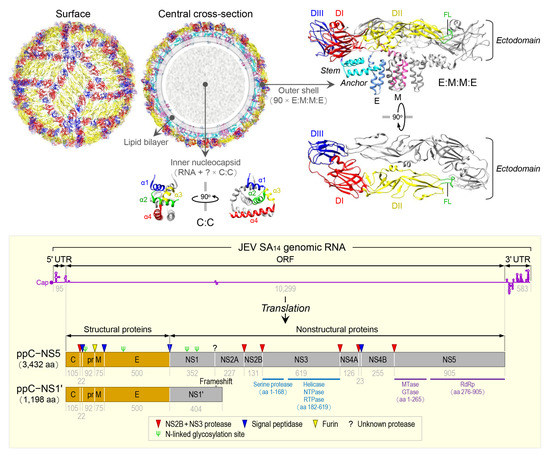
Figure 1. Virion structure, genome organization, and gene expression of JEV. The top panel shows the high-resolution cryo-electron microscopy structure of JEV strain P3 [91][102]. The virion contains multiple copies of three proteins: capsid (C), envelope (E), and membrane (M). The C monomer is a cytosolic protein containing four helices (α1, blue; α2, green; α3, yellow; and α4, red). The E monomer is an integral membrane protein comprising three topologically distinct parts: an N-terminal ectodomain, which has three structural domains (DI, red; DII, yellow; and DIII, blue) with the fusion loop (FL, green) positioned at the distal end of DII; a stem (cyan), which has three non-membrane-spanning helices; and a C-terminal anchor (cornflower blue), which has two membrane-spanning helices. The M monomer is also an integral membrane protein comprising three topologically distinct parts: an N-terminal extension containing an unstructured peptide fragment (pink), a non-membrane-spanning helix (hot pink), and a C-terminal anchor containing two membrane-spanning helices (deep pink). The bottom panel depicts the genome organization and gene expression of JEV strain SA14 [92][103]. The genome is a capped but unpolyadenylated plus-strand RNA, with a single long open reading frame (ORF) flanked by short, highly structured 5’ and 3’ untranslated regions (UTRs). The ORF is translated into two polyprotein precursors, both of which are cleaved by viral and cellular proteases, as indicated, to produce three structural (orange) and seven nonstructural (gray) proteins. Of these proteins, two are multifunctional enzymes: First, NS3 has serine protease, helicase, NTPase, and RTPase activities. Second, NS5 has MTase, GTase, and RdRp activities. Four N-linked glycosylation sites are marked, one in the pr portion of prM, one in E, and two in NS1/1’.