Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Alfred Zheng and Version 2 by Alfred Zheng.
Sulforaphane, derived from broccoli and other members of the Brassica genus, is a phytochemical shown to have anticancer properties. Numerous studies have shown that sulforaphane prevents the development and progression of prostatic tumors. Sulforaphane has been proposed to prevent prostate carcinogenesis by disrupting the Androgen receptor (AR) signaling pathway. Sulforaphane interacts with the promoter region of the AR gene, preventing the transcription of ARs.
- sulforaphane
- prostate cancer
- broccoli
- myrosinase
1. Introduction
The gland known as the prostate is located in the male reproductive system just behind the bladder and surrounds the urethra. The unrestricted proliferation of cells of the prostate gland results in prostate cancer [1]. Prostate carcinoma is one of the most prevalent forms of cancer in men globally, accounting for about 1.4 million new cases and 375,000 mortality per year worldwide [2]. Factors that increase the risk of developing prostate cancer include: age, genetics and lifestyle habits [3]. Considering how common prostate cancer is, the scientific community has intensified efforts in the search for novel therapeutics from naturally occurring compounds capable of preventing, inhibiting or reversing tumor development. Plants have been extensively screened for phytochemicals with anticancer properties; one such phytochemical is sulforaphane [4].
Sulforaphane is a small chemical compound found in cruciferous vegetables of the Brassica genus (broccoli, broccoli sprouts, kale, cabbage, Brussel sprouts and cauliflower). It is produced when the vegetable is chopped, chewed, boiled or disrupted, causing a plant enzyme myrosinase (EC 3.2.1.147) to convert a precursor molecule called glucoraphanin into sulforaphane.
In the 1990s, sulforaphane was isolated from broccoli for the first time and shown to possess anticancer properties by researchers at Johns Hopkins School of Medicine [5][6]. Subsequently, there has been a plethora of studies reporting the antineoplastic activity of sulforaphane. Recent studies have demonstrated that sulforaphane can prevent the development of cancer cells and initiate apoptosis in a variety of cancer types, including prostate cancer [7]. This is due to the compound’s ability to target multiple signaling pathways involved in cancer cell growth and survival [8][9][10].
2. Mechanisms of Action
2.1. AR Signaling
Androgen receptor (AR) signaling mediates the initial stages of prostate carcinogenesis [11]. AR is a hormone receptor and transcription factor. Binding of androgen (such as testosterone) to AR activates AR. The activated AR migrates to the nucleus where it upregulates the transcription of the genes of proteins such as B-cell lymphoma-extra-large (Bcl-XL) and Hypoxia-inducible factor (HIF-1α) [12][13]. Bcl-XL is a protein that suppresses apoptosis and thus promotes the survival and expansion of prostate cancer cells [14]. HIF-1α upregulates the transcription of hexokinase (HK) and pyruvate kinase (PK); over-expression of these enzymes reprograms the metabolism of cells to solely aerobic glycolysis [15]. This is a hallmark of cancer cells known as the Warburg effect [16]. Hence, HIF-1α promotes the development and multiplication of prostate cancer cells via glycolytic metabolism. Sulforaphane has been proposed to prevent prostate carcinogenesis by disrupting the AR signaling pathway. Sulforaphane interacts with the promoter region of the AR gene, preventing the transcription of ARs. This significantly reduces the synthesis of ARs, with no ARs present in the cell surface, androgens cannot bind to ARs to initiate the AR signaling cascade [17] (Figure 1). More so, sulforaphane has been shown to suppress HIF-1α [18]. Sulforaphane binds to HIF-1α and distorts its structure; the distorted HIF-1α loses its function and it is subsequently degraded (Figure 1).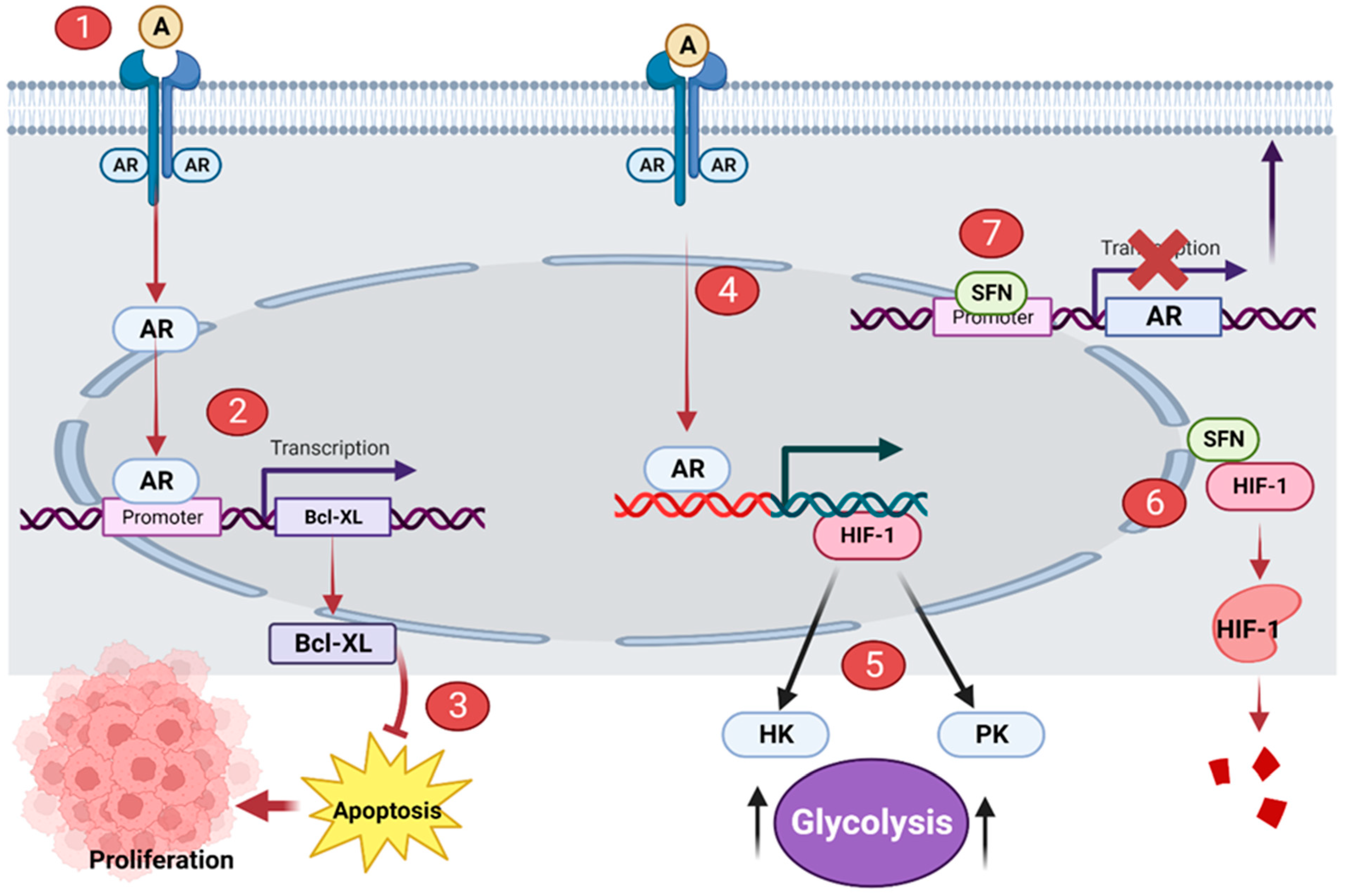
Figure 1. Sulforaphane (SFN) disrupts AR signaling in prostatic tumor cells. (1) Androgen (A) binds to AR. (2) Activated AR upregulates the transcription of Bcl-XL. (3) Bcl-XL suppresses apoptosis and promotes the proliferation of prostate cancer cells. (4) Activated AR upregulates the transcription of HIF-1. (5) HIF-1 promotes the overexpression of HK and PK; these enzymes initiate the Warburg effect. (6) SFN distorts the structure of HIF-1, the distorted protein is subsequently degraded. (7) SFN binds to the promoter region of the AR gene, interrupting its transcription and synthesis. This illustration was made with Biorender.com (accessed on 7 March 2023).
2.2. Induction of Apoptosis
Apoptosis is a natural occurrence through which the number of cells in tissues are regulated. Cancer develops when apoptosis fails; thus, cancerous tissues often suppress apoptosis in cells [19]. Apoptosis can be induced by the activity of the ubiquitin proteasome system (UPS). The UPS involves two processes: ubiquitination and 26S proteasome-mediated degradation. Improperly folded or damaged proteins are marked by ubiquitin, and then recognized and degraded by 26S proteasome [20]. The 26S proteasome has two subunits: a 20S barrel-shaped catalytic core and a 19S regulatory particle. Deubiquitinating enzymes (DUBs) are attached to the 19S regulatory particle to prevent erroneous degradation of cellular proteins. DUBs remove ubiquitin from poly-ubiquitinated protein, preventing its degradation by the proteasome [21]. Tumor tissues often over-express DUBs such as USP14 and UCHL5, thus preventing degradation of proteins and apoptosis, ultimately resulting in the survival and proliferation of cancerous tissues. Sulforaphane has been shown to inhibit the two proteasomal cysteine DUBs, USP14 and UCHL5, in prostate cancer cells [22]. Sulforaphane interacts with USP14 and UCHL5 and suppresses their activity. This promotes increased degradation and induces apoptosis of cells of prostate tumor tissue (Figure 2).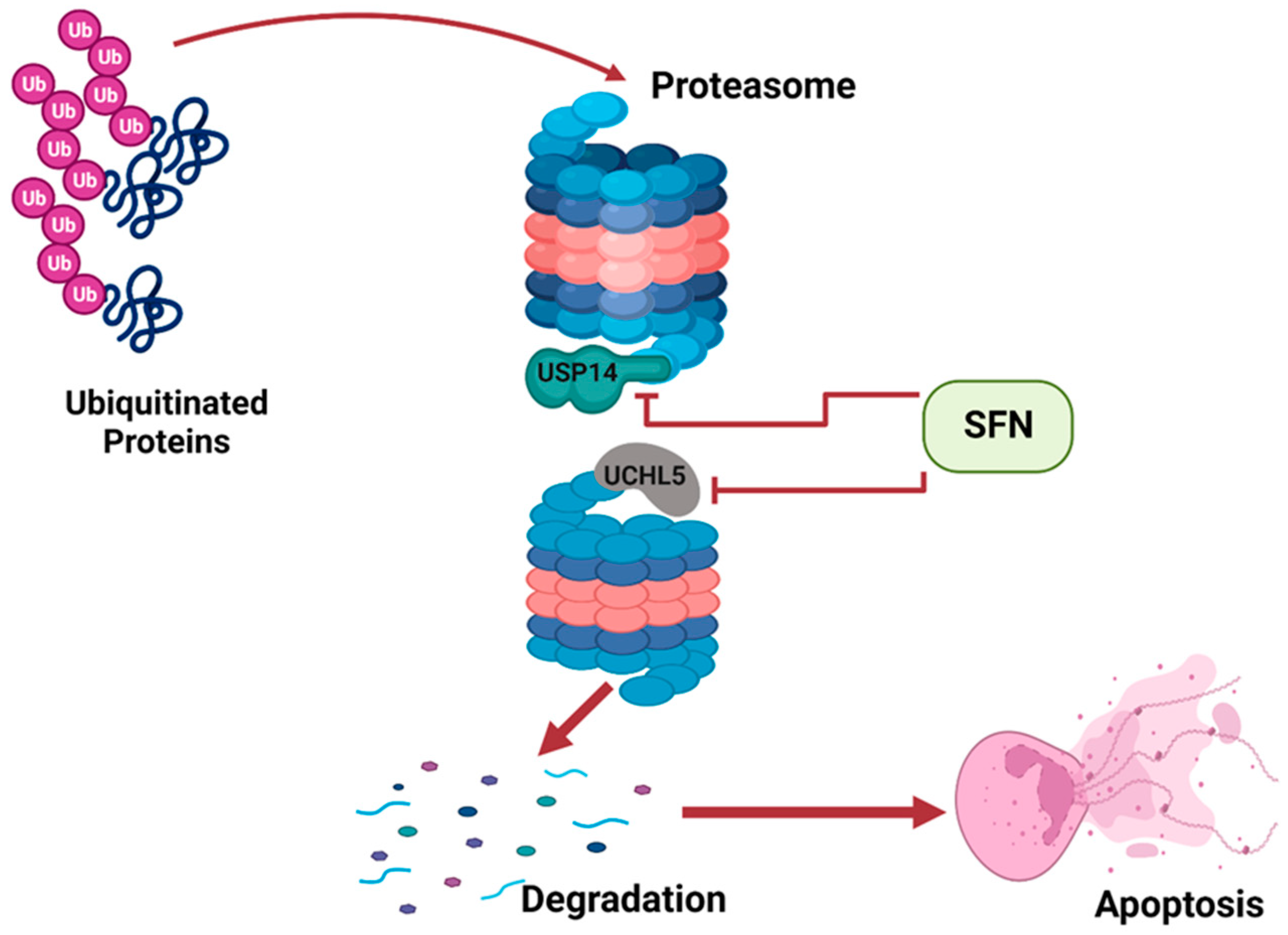
Figure 2.
SFN inhibits DUBs of the UPS to induce apoptosis. This illustration was created with Biorender.com (accessed on 7 March 2023).
2.3. DNA Damage
It has been reported that sulforaphane causes double-stranded DNA breaks and then prevents the repair of these breaks in human prostate cancer cells [23][24]. When DNA damage occurs via a double-stranded break, the repair process involves a complex of various nucleotide excision repair proteins. The combined action of MRN and CtIP proteins holds each pair of single DNA strands in place. RPA, BRCA and XPA work together to form a Holliday junction and a primer at the point of repair. Eventually, DNA synthesis is initiated and the damage is repaired [25]. However, in prostate cancer cells, sulforaphane inhibits XPA protein, an important protein involved in nucleotide excision repair. This disrupts and prevents the repair process; multiple double-stranded DNA breaks accumulate in the cell until the cell is destroyed by apoptosis [24] (Figure 3).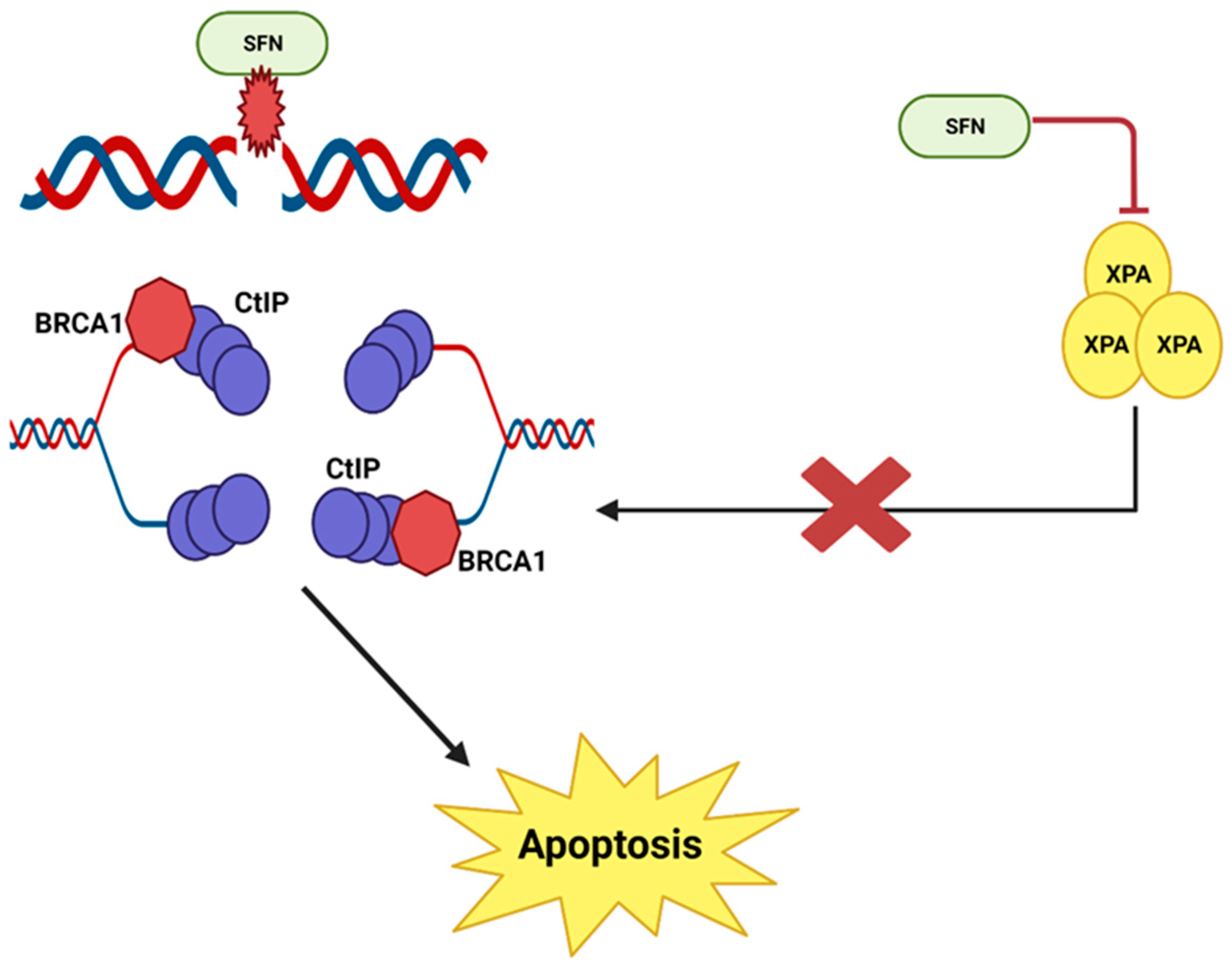
Figure 3.
SFN causes DNA damage and prevents DNA repair in prostate cancer cells. This illustration was made with Biorender.com (accessed on 7 March 2023).
2.4. Upregulation of Protective Enzymes
Sulforaphane protects against prostate carcinogenesis by upregulating the transcription of carcinogen-detoxifying enzymes (Phase 2 enzymes). Sulforaphane binds to Keap1 in the cytoplasm and disrupts its orientation. This disruption releases Nuclear factor erythroid 2 (Nrf2). Nrf2 is transported to the nucleus, where it binds to antioxidant response element (ARE); this leads to the increased transcription of Phase 2 detoxifying enzymes such as quinone 1, NAD(P)H dehydrogenase and heme oxygenase 1. These enzymes enhance cellular defenses and prevent the initiation of carcinogenesis [26][27] (Figure 4).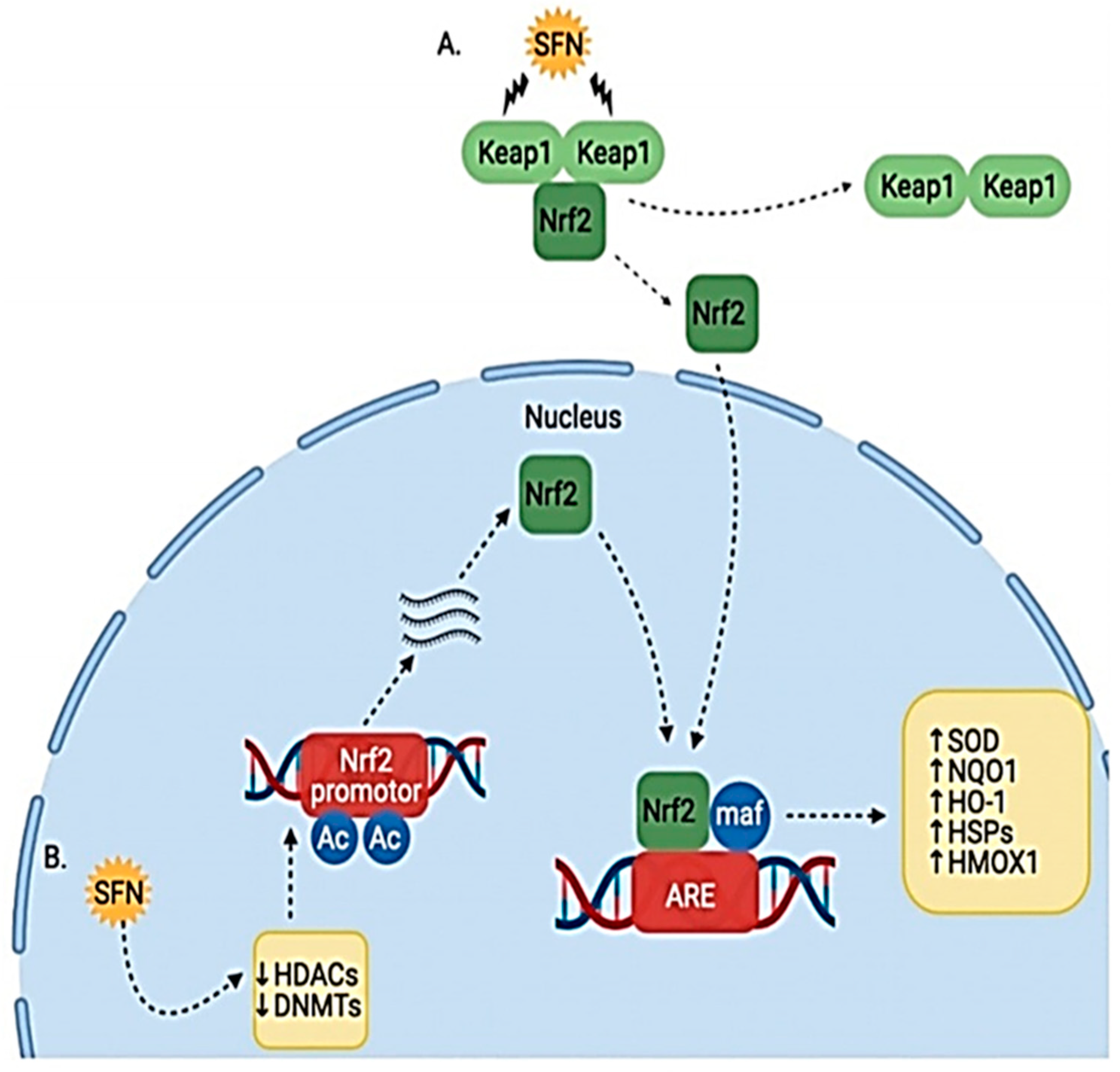
Figure 4. Effect of SFN on Nrf2/Keap1 pathway. (A) SFN modifies Keap1 cysteine residues, causing release of Nrf2, which allows it to translocate to the nucleus. (B) SFN also induces epigenetic modulation of HDACs and DNMTs, causing increased Nrf2 transcription and translation [26].
2.5. Autophagy
Autophagy is a process by which the cell maintains homeostasis by recycling old and damaged cytoplasmic components such as proteins and organelles [28]. The process involves the formation of membranous vacuoles (autophagosomes), which engulf the cytoplasmic components marked for recycling. Autophagosomes fuses with lysosomes, and the lysosomal enzymes degrade the content of the vacuoles [29]. Researchers have shown that autophagy plays a complex role in the development and progression of cancer cells [30]. In the early stages of prostate cancer, sulforaphane induces autophagy by upregulating the transcription of microtubule-associated protein 1 light chain 3 (LC3), an essential protein for the formation of autophagosomes [31]. This induction of autophagy results in cytoprotective effects on prostate cells and the suppression of further progression of prostatic tumors, as damaged and abnormal cell organelles are rapidly degraded [32]. However, in the later stages of the carcinoma, autophagy promotes the survival of cancer cells by shielding them from the effects of stress and therapy. Thus, autophagy inhibitors (such as chloroquine) have been proposed as an adjuvant to sulforaphane for advanced cases of prostate cancer [33].References
- Leslie, S.W.; Soon-Sutton, T.L.; R I, A.; Sajjad, H.; Siref, L.E. Prostate Cancer. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022.
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249.
- Kalish, L.A.; McDougal, W.S.; McKinlay, J.B. Family history and the risk of prostate cancer. Urology 2000, 56, 803–806.
- Brooks, J.D.; Paton, V.G.; Vidanes, G. Potent induction of phase 2 enzymes in human prostate cells by sulforaphane. Cancer Epidemiol. Biomark. Prev. 2001, 10, 949–954.
- Zhang, Y.; Talalay, P.; Cho, C.G.; Posner, G.H. A major inducer of anticarcinogenic protective enzymes from broccoli: Isolation and elucidation of structure. Proc. Natl. Acad. Sci. USA 1992, 89, 2399–2403.
- Zhang, Y.; Kensler, T.W.; Cho, C.G.; Posner, G.H.; Talalay, P. Anticarcinogenic activities of sulforaphane and structurally related synthetic norbornyl isothiocyanates. Proc. Natl. Acad. Sci. USA 1994, 91, 3147–3150.
- Na, G.; He, C.; Zhang, S.; Tian, S.; Bao, Y.; Shan, Y. Dietary Isothiocyanates: Novel Insights into the Potential for Cancer Prevention and Therapy. Int. J. Mol. Sci. 2023, 24, 1962.
- Lewinska, A.; Bednarz, D.; Adamczyk-Grochala, J.; Wnuk, M. Phytochemical-induced nucleolar stress results in the inhibition of breast cancer cell proliferation. Redox Biol. 2017, 12, 469–482.
- Dong, Q.Q.; Wang, Q.T.; Wang, L.; Jiang, Y.X.; Liu, M.L.; Hu, H.J.; Liu, Y.; Zhou, H.; He, H.P.; Zhang, T.C.; et al. SMYD3-associated pathway is involved in the anti-tumor effects of sulforaphane on gastric carcinoma cells. Food Sci. Biotechnol. 2018, 27, 1165–1173.
- Yin, L.; Xiao, X.; Georgikou, C.; Luo, Y.; Liu, L.; Gladkich, J.; Gross, W.; Herr, I. Sulforaphane Induces miR135b-5p and Its Target Gene, RASAL2, thereby Inhibiting the Progression of Pancreatic Cancer. Mol. Ther. Oncolytics 2019, 14, 74–81.
- Zhou, Y.; Bolton, E.C.; Jones, J.O. Androgens and androgen receptor signaling in prostate tumorigenesis. J. Mol. Endocrinol. 2015, 54, R15–R29.
- Tan, M.H.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E.L. Androgen receptor: Structure, role in prostate cancer and drug discovery. Acta Pharmacol. Sinica. 2015, 36, 3–23.
- Mabjeesh, N.J.; Willard, M.T.; Frederickson, C.E.; Zhong, H.; Simons, J.W. Androgens stimulate hypoxia-inducible factor 1 activation via autocrine loop of tyrosine kinase receptor/phosphatidylinositol 3’-kinase/protein kinase B in prostate cancer cells. Clin. Cancer Res. 2003, 9, 2416–2425.
- Michels, J.; Kepp, O.; Senovilla, L.; Lissa, D.; Castedo, M.; Kroemer, G.; Galluzzi, L. Functions of BCL-X L at the Interface between Cell Death and Metabolism. Int. J. Cell Biol. 2013, 2013, 705294.
- Jun, J.C.; Rathore, A.; Younas, H.; Gilkes, D.; Polotsky, V.Y. Hypoxia-Inducible Factors and Cancer. Curr. Sleep Med. Rep. 2017, 3, 1–10.
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033.
- Carrasco-Pozo, C.; Tan, K.N.; Rodriguez, T.; Avery, V.M. The Molecular Effects of Sulforaphane and Capsaicin on Metabolism upon Androgen and Tip60 Activation of Androgen Receptor. Int. J. Mol. Sci. 2019, 20, 5384.
- Yao, H.; Wang, H.; Zhang, Z.; Jiang, B.H.; Luo, J.; Shi, X. Sulforaphane inhibited expression of hypoxia-inducible factor-1alpha in human tongue squamous cancer cells and prostate cancer cells. Int. J. Cancer 2008, 123, 1255–1261.
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348.
- Lee, B.H.; Lee, M.J.; Park, S.; Oh, D.C.; Elsasser, S.; Chen, P.C.; Gartner, C.; Dimova, N.; Hanna, J.; Gygi, S.P.; et al. Enhancement of proteasome activity by a small-molecule inhibitor of USP14. Nature 2010, 467, 179–184.
- Liao, Y.; Liu, N.; Hua, X.; Cai, J.; Xia, X.; Wang, X.; Huang, H.; Liu, J. Proteasome-associated deubiquitinase ubiquitin-specific protease 14 regulates prostate cancer proliferation by deubiquitinating and stabilizing androgen receptor. Cell Death Dis. 2017, 8, e2585.
- Ahmed, Z.S.O.; Li, X.; Li, F.; Cheaito, H.A.; Patel, K.; Mosallam, E.M.; Elbargeesy, G.A.E.H.; Dou, Q.P. Computational and biochemical studies of isothiocyanates as inhibitors of proteasomal cysteine deubiquitinases in human cancer cells. J. Cell. Biochem. 2018, 119, 9006–9016.
- Hać, A.; Brokowska, J.; Rintz, E.; Bartkowski, M.; Węgrzyn, G.; Herman-Antosiewicz, A. Mechanism of selective anticancer activity of isothiocyanates relies on differences in DNA damage repair between cancer and healthy cells. Eur. J. Nutr. 2020, 59, 1421–1432.
- Sekine-Suzuki, E.; Yu, D.; Kubota, N.; Okayasu, R.; Anzai, K. Sulforaphane induces DNA double strand breaks predominantly repaired by homologous recombination pathway in human cancer cells. Biochem. Biophys. Res. Commun. 2008, 377, 341–345.
- Brochier, C.; Langley, B. Chromatin modifications associated with DNA double-strand breaks repair as potential targets for neurological diseases. Neurother. J. Am. Soc. Exp. NeuroTherapeutics 2013, 10, 817–830.
- Kaiser, A.E.; Baniasadi, M.; Giansiracusa, D.; Giansiracusa, M.; Garcia, M.; Fryda, Z.; Wong, T.L.; Bishayee, A. Sulforaphane: A Broccoli Bioactive Phytocompound with Cancer Preventive Potential. Cancers 2021, 13, 4796.
- Fahey, J.W.; Talalay, P. Antioxidant functions of sulforaphane: A potent inducer of Phase II detoxication enzymes. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1999, 37, 973–979.
- Lum, J.J.; DeBerardinis, R.J.; Thompson, C.B. Autophagy in metazoans: Cell survival in the land of plenty. Nat. Rev. Mol. Cell Biol. 2005, 6, 439–448.
- Seglen, P.O.; Bohley, P. Autophagy and other vacuolar protein degradation mechanisms. Experientia 1992, 48, 158–172.
- Ogier-Denis, E.; Codogno, P. Autophagy: A barrier or an adaptive response to cancer. Biochim. Et Biophys. Acta 2003, 1603, 113–128.
- Herman-Antosiewicz, A.; Johnson, D.E.; Singh, S.V. Sulforaphane causes autophagy to inhibit release of cytochrome C and apoptosis in human prostate cancer cells. Cancer Res. 2006, 66, 5828–5835.
- Paglin, S.; Hollister, T.; Delohery, T.; Hackett, N.; McMahill, M.; Sphicas, E.; Domingo, D.; Yahalom, J. A novel response of cancer cells to radiation involves autophagy and formation of acidic vesicles. Cancer Res. 2001, 61, 439–444.
- Vyas, A.R.; Hahm, E.R.; Arlotti, J.A.; Watkins, S.; Stolz, D.B.; Desai, D.; Amin, S.; Singh, S.V. Chemoprevention of prostate cancer by d,l-sulforaphane is augmented by pharmacological inhibition of autophagy. Cancer Res. 2013, 73, 5985–5995.
More