Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Marwa Omar and Version 2 by Peter Tang.
Neutrophils are the key players in the innate immune system, being weaponized with numerous strategies to eliminate pathogens. The production of extracellular traps is one of the effector mechanisms operated by neutrophils in a process called NETosis. Neutrophil extracellular traps (NETs) are complex webs of extracellular DNA studded with histones and cytoplasmic granular proteins.
- extracellular traps
- protozoa
- innate immunity
- NETosis
- helminths
- neutrophils
- DNA
1. Introduction
Parasitic helminths (metazoa) and protozoa are vastly diverse groups of eukaryotic organisms that cause various diseases in humans and animals. Parasitic infections pose a major public health concern in developing countries, mainly in tropical and subtropical communities [1]. Parasites can cause persistent infection due to their ability to resist immune-mediated expulsion by modulating the host’s immune response [2][3][4][2,3,4]. The intricate interaction between the parasite and the host is critically important when understanding the pathophysiology of the diseases they cause [5].
Innate immunity is an ancient form of the host’s defensive mechanisms against invasive pathogens. This type of immunity has evolved to protect the host against different infectious agents, including bacteria, fungi, viruses, and parasites [6]. The components of the innate machinery include physical and anatomical barriers, along with humoral and cellular factors [7][8][7,8]. Neutrophils, monocytes, and macrophages are among the professional phagocytic cells participating in both phagocytosis and inflammatory processes [9][10][9,10].
The generation of extracellular traps (ETs) has been recognized as a novel mechanism of the innate immune response against infections. These traps are expelled to facilitate the immobilization and killing of invading microorganisms in the extracellular environment [11]. ETs can be generated by several leukocytes, including neutrophils, eosinophils, basophils, and mast cells [12]. In addition, activated T cells, B cells, and monocytes are likely to release mitochondrial DNA (mtDNA), forming extracellular web-like structures. Regardless of the type of cells from which they originate, the traps share a common feature: they consist of nuclear or mtDNA as a backbone, with embedded antimicrobial peptides, histones, and cell-specific proteases [13].
2. Neutrophils and Neutrophil Extracellular Traps (NETs)
2.1. Neutrophils
Neutrophils, or poly-morphonuclear (PMN) leukocytes, are the most abundant cell type circulating in human blood. They are considered the foot soldiers of the immune system and the first comers to the sites of infection or inflammation [14][15][14,15]. Neutrophils play a pivotal role in developing innate and adaptive immune responses [16]. They employ different strategies to combat pathogens, including phagocytosis [17][18][17,18], degranulation [19][20][19,20], and the formation of neutrophil extracellular traps (NETs) [21].
2.2. NETs and NETosis
The original description of NET production was first introduced by Brinkmann et al. (2004) as a new defensive mechanism consisting of the ejection of intracellular material in the form of web-like elements into the surrounding extracellular medium. The authors discovered that when stimulated with phorbol myristate acetate (PMA), lipopolysaccharide (LPS), and interleukin 8 (IL-8), neutrophils release nuclear DNA fibers after exposure to Gram-positive or Gram-negative bacteria [21]. NETs are composed of a backbone of DNA decorated with histones and laced with several cytoplasmic antimicrobial granular proteins [21][22][23][21,22,23]. The granular effector molecules include myeloperoxidase (MPO), neutrophil elastase (NE), lactoferrin, cathepsins, pentraxin, gelatinase, peptidoglycan recognition protein (PRP), calprotectin, bacterial permeability-increasing protein (BPIP), and other leukocyte peptides [24][25][26][24,25,26]. The fibrous DNA strands in the NETs are punctuated with globular protein domains in large aggregates ranging from ~25 nm up to 50 nm in size. The chromatin constitutes the backbone of these structures, as NETs can be degraded by DNases, but not by proteases [27].
The whole process of NET production is called NETosis, a phenomenon first described in the work of Takei et al. (1996) [28], based on the term ‘NET’ [22] and the Greek suffix ‘-osis’ [29]. It is a novel form of cell death that is distinct from autophagy or necrosis, as it is independent of the caspase pathway, and there is no phosphatidylserine exposition on the cell surface [22]. Furthermore, NETosis is not associated with DNA fragmentation, as is the case in apoptosis, but rather with the release of NETs after the disruption of the plasma membrane and nuclear envelope. The sticky DNA-rich networks not only entrap invading pathogens, but eventually kill them [30].
Various reports have defined NETosis as an NADPH oxidase (NOX)-dependent mechanism [21][31][21,31]. Under stimulation, neutrophils generate reactive oxygen species (ROS) by NADPH oxidase-2 (NOX-2) [32]. ROS modulate the granular enzyme MPO, and both are required for the release of NE from granules and its translocation to the nucleus [33][34][33,34]. In the nucleus, NE contributes to chromatin decondensation by the proteolysis of histones. Such events result in the nuclear extrusion of DNA and NET production [22][32][22,32]. Yet, other studies have described a (NOX)-independent NET formation process, which is facilitated through calcium influx and mitochondrial ROS production [35][36][35,36]. Hence, this type of NET expulsion can be called “mitochondria-dependent NETosis” [37].
2.3. Mechanisms of NET Formation
There are at least three different mechanisms by which NETs are formed: classical or suicidal NETosis, the noncanonical pathway, and vital NETosis. The lytic or suicidal type usually occurs slowly (2−4 h) and involves the rupture of the neutrophil plasma membrane. On the other hand, nonlytic or vital NETosis occurs rapidly (within minutes) and does not involve disruption to the plasma membrane or cell lysis [38]. During vital NETosis, neutrophils are still capable of functions such as migration, degranulation, and phagocytosis while casting their sticky filaments [39]. The noncanonical pathway is a new pathway of lethal NETosis that has recently been described. This sort suppresses bacterial residence in the neutrophil cytosol and prevents in vivo microbial dissemination [40].
2.4. Microbial Triggers of NETosis
Multiple stimuli promote the release of NETs. These include the interaction of neutrophils with other immune cells (platelets) after activation with cytokines (IL-8) that help to entrap and eliminate pathogens [41]. Additionally, neutrophils induce vital NETosis when exposed either in vitro or in vivo to whole microorganisms or their proteins [39]. The microbicidal effects of NETs have been confirmed in several human and animal models with bacterial [21], viral [42], and fungal infections [43].
NETs entrap many types of bacterial pathogens to prevent their spread. A variety of Gram-positive and Gram-negative bacteria have been shown to induce NET expulsion. Infection with the swine pathogen Streptococcus suis leads to the signaling of NET formation pathways in a NOX-dependent approach [44]. Additionally, the Gram-negative bacterium Klebsiella pneumoniae proved to be a good inducer of NETosis in a mouse lung infection model [45]. Yet, some bacteria have developed strategies to resist their capture or elimination by the NET structures. Staphylococcus aureus produces different enzymes to interfere with the antimicrobial properties of NETs, such as a nuclease (Nuc), a DNA binding protein, and the extracellular adherence protein (Eap) [46]. These enzymes facilitate the escape from the NET filaments, delay bacteria clearance, and increase the mortality caused by the infection [47]. Besides the reported role of bacteria in triggering NETosis, increasing evidence indicates that viruses can also promote NET formation, which may either promote or prevent viral-induced pathology [48]. In the case of Human Immunodeficiency Virus-1 (HIV-1), NETs promote pathogen clearance through the actions of histones and MPO [49]. Recently, Veras et al. [50] demonstrated an increase in the quantity of the NET components (DNA-MPO complex) in the plasma, tracheal aspirate, and autopsy lung tissues of COVID-19 patients. The study highlighted a possible detrimental role of NETs in the pathophysiology of COVID-19.
Neutrophils and lymphocytes are closely related to the pathophysiology of several inflammatory disorders. The neutrophil-to-lymphocyte ratio (NLR) is a biomarker that conjugates two arms of the immune system: the innate immune response, mainly due to neutrophils, and adaptive immunity, promoted by lymphocytes. This ratio is characterized by an increase in neutrophils and a decline in lymphocytes [51]. In their research, Regolo et al. (2022) tried to assess the prognostic value of (NLR) as a predictor of the outcome of COVID-19 patients. According to them, patients with higher NLR showed a higher risk of intra-hospital mortality and disease progression [52].
The filamentous fungus Candida albicans (C. albicans) is the most widely discussed fungal pathogen in the field of NET release. Following the interaction of neutrophils with C.albicans, the granular protein (NE) is released into the cytoplasm to initiate the production of NETs, which have been shown to kill both yeast and hyphal forms of the fungus [43]. Yet, C. albicans has its unique way of resisting NETs. It potentially modulates the formation of NETs by arresting their proteinaceous components, including elastase, MPO, and histones. Additionally, adhesins on the fungal surface can adsorb the NETs proteins and increase the pathogen’s potency in host tissue destruction [53].
3. NETosis in Parasitic Infections
The functional responses of neutrophils to parasitic infections continue to be uncovered. It has been suggested that the size of the stimulating particle is one of the factors driving the decision of neutrophils to generate NETs instead of phagocytosis. While large particles, such as parasites, can induce the formation of NETs, small particles, such as bacteria, yeast, or viruses, can be eliminated by phagocytosis [54][55][54,55]. Knowledge is not yet abundant about the involvement of NETs in the host’s innate response against metazoan and protozoan parasites [5] (Figure 1).
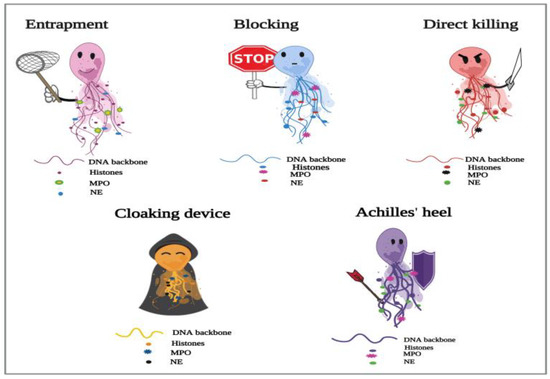
Figure 1. Proposed depiction of the functional responses of NETs to parasitic infections. In the process of NETosis, neutrophils release complex DNA webs, in which actions extend from parasitic entrapment to blocking the worm development and effector killing. Further, NETs could serve as a “cloaking device” to control the spread of infection. ThWe researchers aalso have created the “Achilles’ heel of NETs” to frame the several reported limitations in their actions in answer to different parasites.
3.1. NETosis and Metazoan Infections
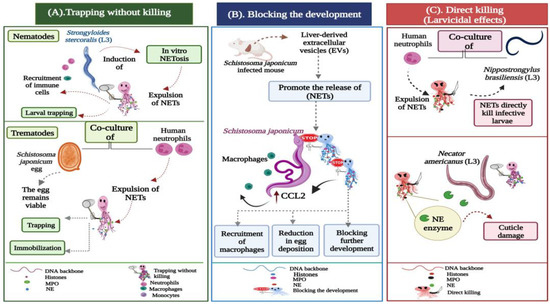
Figure 2. Potential actions operated by NETs during metazoan infections. (A) Trapping without killing: Strongyloides stercoralis large and motile larvae are potent inducers of in vitro NETosis. During the infection, the DNA threads ensnare larvae to enhance their killing by the recruited leukocytes, neutrophils, and macrophages. In trematode-triggered NETosis, the NET filaments capture the egg stages of Schistosoma japonicum. NETs hinder the mobility of the eggs, which still remain viable and intact within the fibrous traps. (B) Blocking the development: Extracellular vesicles (EVs) isolated from the liver of Schistosoma japonicum-infected mice initiate the expulsion of NETs, which block worm development, resulting in a significant reduction in egg deposition and associated fibrosis. The extruded NETs also promote chemokine ligand 2 (CCL2) expression in macrophages, which adhere to the surface of Schistosoma japonicum worms, inhibiting their further development. (C) Direct larvicidal effects: The direct toxicity of neutrophils is distinctly exhibited against the infective stages of hookworms. The skin-penetrating third-stage larvae (L3) of Nippostrongylus brasiliensis and Necator americanus become mechanically trapped by NETs, which directly participate in the larval killing using the neutrophil elastase (NE) enzyme, which induces cuticle damage to the hookworm larvae.
-
Trapping without killing;
-
Blocking the development;
-
Direct killing (larvicidal) effects.