Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Livia Marques Casanova and Version 2 by Rita Xu.
Allelochemicals are toxic secondary metabolites produced by plants, microalgae, bacteria, and fungi that influence other organisms. The bioactivity of allelochemicals and their toxic or beneficial effects have been the focus of research in medicine and agriculture, and for their anticancer and antimicrobial properties. Microalgae are the source of a remarkable diversity of biologically active compounds, which can be involved in allelopathic interactions. The main chemical classes of microalgal allelochemicals are alkaloids, fatty acids and derivatives, polyketides, peptides, phenolics, and terpenoids. In the environment, these molecules are secreted by microalgae for communication, defence, and adaptation purposes.
- allelopathy
- allelochemical
- microalgae
1. Introduction
Allelopathy is an ecological phenomenon in which allelochemicals are produced under certain environmental conditions. The term ‘allelopathy’ was introduced by Molisch in 1937 and originated from the Greek words ‘allelon’ (mutual) and “pathos” (harm). The term means one organism influences others through a chemical mode [1].
Allelochemicals are toxic secondary metabolites produced by plants, microalgae, bacteria, and fungi that influence the development of other organisms. These substances can be harmful to humans, other microalgae, and organisms, including phytoplankton, animals, and submerged macrophytes. Photosynthesis and enzyme activity inhibition, damage to cell structures, and other physiological alterations such as inhibition of respiration, protein synthesis, and gene expression have been observed [2][3][2,3].
Allelopathy is much studied in plants, but allelochemicals started to be reviewed in microalgae by Inderjit and Dakshini in 1994 [2][4][2,4]. Microalgae are a widely diversified group of prokaryotic and eukaryotic photosynthetic microorganisms. More than 50,000 microalgae species are distributed in rivers, lakes, oceans, and even in terrestrial ecosystems. Cyanobacteria are prokaryotic microalgae in which membrane-bound organelles such as plastids and mitochondria are missing, while eukaryotic microalgae rely on these structures to control cellular functions [5][6][5,6]. Many common microalgae taxa are included in the latter group, such as Chlorophyta (green algae) and Bacillariophyta (diatoms) [5][6][5,6]. Pyrrophyta (dinoflagellates) are mostly microplankton, they are eukaryotic and live in aquatic environments, be it salt, brackish or fresh water. Around half of all dinoflagellates are photosynthetic, while the other half are heterotrophic; some live as mixotrophic and obligate symbionts/parasites. They are usually reported as microalgae; indeed, the most toxic microalgae are dinoflagellates [7] The non-algal dinoflagellates are primarily predators of other microorganisms or parasites of animals or giant protists [8].
One of the fields of study for allelochemicals in microalgae began with harmful algal blooms (HABs). HABs are a consequence of an ecosystem imbalance in marine and freshwater systems worldwide [9]. HABs affect water usage, including fisheries, water resources, and recruitment. The poisoning caused by the allelochemicals affects different organisms and leads to economic losses [10]. The use of allelochemicals as biocontrol agents has been suggested as a means of improving microalgae cultivation and controlling contamination [11]. An example of a mode of action is the generation of reactive oxygen species (ROS). Nostoc spongiaeforme produces a pigment named nostocine A; this pigment is highly cytotoxic for several microalgae. The compound accelerates ROS formation in Chlorella reinhardtii. Nostocine A penetrates the target and is reduced by NADPH; when oxygen levels are high, reduced nostocine A is oxidated, generating superoxide radical anion (O2−) that may cause cytotoxicity [12].
The bioactivity of allelochemicals and their toxic or beneficial effects have also been the focus of research in medicine and agriculture [13], and for their anticancer [14] and antimicrobial properties (Figure 1) [15].
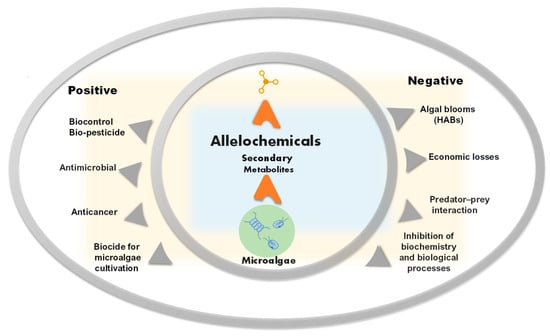
Figure 1. Schematic representation of positive and negative aspects of allelochemicals from microalgae.
2. Allelopathy in Microalgae
As mentioned previously, the term allelopathy was originally used to describe biomolecules produced by plants. However, it is now openly used with other organisms and has been studied in microalgae [16][17][19,20]. Microalgae organisms produce numerous secondary metabolites, including fatty acids, alkaloids, amino acids, and peptides, which are investigated and characterized as natural products by chemists and described as novel bio compounds. In the environment, these molecules are secreted for communication, defence, and adaptation purposes [18][21]. More knowledge is needed in order to understand the complexity of these bioactive chemicals and their interaction with and effect on ecosystems [16][19][19,22]. A recent review paper showed that Microcystis, Nodularia, Karenia, Alexandrium (A. catenella, A. fundyense, and others species), Skeletonema, Chlorella (C. vulgaris, C. sorokiniana, C. ellipsoidea), Chlamydomonas, and Dunaliella displayed allelopathic activity against other microalgae and cyanobacteria [20][23]. One strategy to study the allelopathic effects of different microalgae agents is to analyse the biochemicals present in mono- and co-cultures [21][24]. Wang et al. [21][24] observed that the dinoflagellate Scrippsiella acuminata had its growth significantly inhibited when in co-cultivation and/or in the presence of exudates (cell-free and sonicated-cell filtrates) from three diatoms (Chaetoceros curvisetus, Phaeodactylum tricornutum and Skeletonema dohrnii), suggesting the diatoms developed significant competitive advantage over other antagonists. Furthermore, the authors detected several types of volatile organic compounds (VOCs) in the sonicated cell-free filtrates. Polyunsaturated aldehydes (PUAs) were found in all three diatoms that showed inhibitory effects on the growth of S. acuminata and the diatoms themselves. Generally, the production of microalgae VOCs is influenced by environmental conditions, such as nutrient availability, temperature, and light incidence [22][23][25,26]. Studies show that allelochemicals of microalgae have either a positive or negative impact on other organisms. Those which can cause inhibitory effects are suitable for application as biopesticides, and as a new source of antimicrobials or bio-herbicide agents [16][20][19,23], especially because these biocompounds present increased target specificity [24][27]. The liberation of species-specific allelochemicals is desired in agriculture and is an important focus for future studies because they can be responsible for modulating the ecosystem under favourable conditions to an allelopathic microalgae. An example of this was observed in a study where, by alternating the nutrients conditions and balance of nitrogen and phosphorus sources, the allelopathic interaction between strains of Anabaena spp. and Microcystis spp. seemed to control their relative abundance and their dominance in that habitat [25][28]. An interesting point to remark upon here is that one of several new applications of microalgae is as a bioenergy source and an alternative to fossil fuels [26][27][28][29,30,31]. Nevertheless, one of the elements that industry must be alert to is the accumulation of allelopathic molecules released into the water by microalgae themselves. These compounds, when in high concentrations due to high biomass, along with other biomolecules can trigger positive or negative effects on other cells, leading to auto-stimulation or self-inhibition [29][30][31][32,33,34]. Lu and colleagues (2020) described studies that focused on water reuse for microalgae cultivation, and stated that several factors, including allelopathy mechanisms, cause cell damage, aggregation, and programmed cell death (PCD) (negative effects) or, on the other hand, the liberation of growth regulators and removal of growth inhibitors (positive effects). Bacterial agents can have a significant influence on allelopathy affecting microalgae biomass tanks [30][32][33,35]. Some allelopathic molecules have been characterized in the scientific literature as having a role in algal community growth regulation. Satake et al. [33][36] described a 19 membered ring-structured molecule called alexandrolide, which was isolated from dinoflagellate Alexandrium catenella and inhibited diatoms (Skeletonema costatum and Chattonella antiqua) but did not inhibit other dinoflagellates. Marennine, a blue-green polyphenolic pigment produced by diatom Haslea ostrearia, was described as being responsible for inhibiting the growth of other diatom species and self-stimulating the producer species in oyster-ponds [34][37]. By elucidating the properties of these compounds, more studies can be performed to apply these chemicals for industrial and human purposes, such as in additives in agricultural technology [35][38].3. Allelochemicals from Microalgae
Microalgae are the source of a remarkable diversity of biologically active compounds, which can be involved in allelopathic interactions. The main chemical classes of microalgal allelochemicals are: alkaloids, fatty acids and derivatives, polyketides, peptides, phenolics, and terpenoids.3.1. Alkaloids
Alkaloids are a prominent class of allelochemicals with biocidal properties. They consist of nitrogen-containing heterocyclic substances, in which the nitrogen atom is usually derived from an amino acid [36][37][39,40]. Alkaloids have been reported mainly in cyanobacteria and dinoflagellates. In the former, indole alkaloids are the most frequent. For instance, 12-epi-hapalindole E isonitrile (Figure 2A; 1), from Fischerella sp., is an indole alkaloid with activity against green microalgae and other microorganisms [38][41]. Another example is the toxin calothrixin A (Figure 2A; 2) from Calothrix sp [38][41].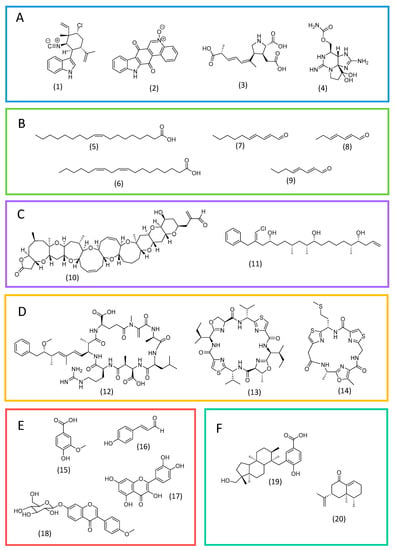
Figure 2. Examples of allelochemicals from microalgae. Alkaloids (A): 12-epi-hapalindole E isonitrile (1), calothrixin A (2), domoic acid (3), and saxitoxin (4). Fatty acids and derivatives (B): oleic acid (5), linoleic acid (6), 2E,4E-decadienal (7), 2E,4E-heptadienal (8), and 2E,4E-octadienal (9). Polyketides (C): brevetoxin A (10) and trichophycin A (11). Peptides (D): microcystin-LR (12), patellamide A (13), and tenuecyclamide C (14). Phenolic substances (E): vanillic acid (15), p-coumaric acid (16), quercetin (17) and ononin (18). Terpenoids (F): comnostin A (19) and eremophilone (20).