Iron is an essential micronutrient because of its importance in the process of erythropoiesis, oxidative metabolism, and cellular immune responses [
1]. Healthy adults contain 4–5 g of iron, which is mostly (65%) found in red blood cell hemoglobin (Hb), and 30–35% is stored in the liver in the form of ferritin. Only 1–2% is in the form of iron-sulfur clusters or heme in the enzymes and multiprotein complexes [
2]. Despite its essential role in the human body [
3], there are no effective means of excreting iron [
4]. Thus, a critical point in iron homeostasis is the regulation of the absorption of dietary iron from the duodenum. The body absorbs 1–2 mg of dietary iron a day, and this intake must be balanced with losses in the form of sloughed intestinal mucosal cells, menstruation, and other blood losses [
5]. Maintaining the balance is very important because free iron is able to generate free radicals through Fenton reaction, and it is highly toxic [
6,
7]. Therefore, organisms have developed sophisticated pathways to import, chaperone, sequester, and export iron in order to maintain an appropriate iron balance [
8]. Any disruption of iron homeostasis leads to either iron deficiency (ID) or iron overload (IO) [
9].
IO leads to adverse manifestations in different tissues (brain, heart, liver, adipose, muscle, pancreas) and it is implicated in the pathogenesis of several metabolic (e.g., type 2 diabetes, non-alcoholic steatohepatitis, atherosclerosis, stroke, etc.) [
12] and neurodegenerative diseases (e.g., Alzheimer’s disease (AD), Parkinson’s disease (PD), multiple sclerosis (MS), etc.) [
13], which could be found more often in elderly.
2. Brain Iron Metabolism
The brain is a very metabolically active organ and accounts for about 20% of the body’s total energy consumption. These high-energy needs must be supported with an adequate supply of iron [
97]. Therefore, iron is the most abundant metal in the brain [
14]. It has an essential role as a co-factor for many physiological processes in the CNS, including oxidative metabolism, myelination, and the biosynthesis of neurotransmitters [
98]. To ensure the normal course of these processes, brain iron levels are tightly regulated [
99].
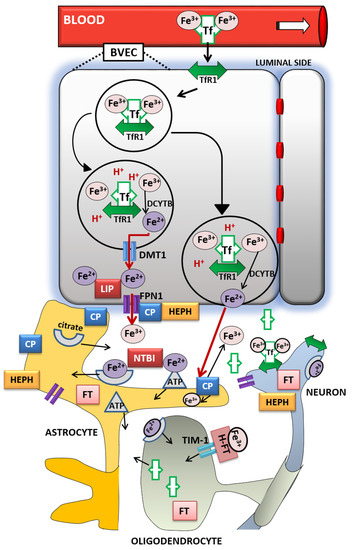
The entry of iron from the blood into the brain is controlled by the blood–brain barrier (BBB) [
100] and to a lesser extent by the blood–cerebrospinal fluid barrier (BCSFB) [
101]. The role of the BBB is to prevent the brain from neurotoxic plasma components and pathogens [
102]. At the same time, it controls chemical composition of the neuronal milieu by regulating the transport of molecules required for normal neuronal functioning [
103]. The BBB is formed by a monolayer of tightly sealed microvascular endothelial cells extending along the vascular tree [
104] and expressing low paracellular and transcellular permeability [
105]. Those endothelial cells are surrounded by basal lamina and astrocytic perivascular end-feet, forming the neurovascular unit [
106]. Tf-bound iron cannot cross the BBB directly and the mechanism of iron transcellular entry into the brain is not entirely clear [
107]. According to the recent models [
108,
109], there are two possible iron transport pathways: transferrin-bound iron (Tf-Fe) and non-transferrin-bound iron (NTBI) [
107] ().
Figure 1. Iron transport inside the brain. A scheme of proposed transferrin-bound and non-transferrin bound iron transport pathways in the brain. Abbreviations: BVEC—blood vascular endothelial cell, Tf—transferrin, TfR1—transferrin receptor 1, Fe3+—ferric iron, Fe2+—ferrous iron, DCYTB—duodenal cytochrome b, DMT1—divalent metal transporter-1, FPN1—ferroportin-1, LIP—labile iron pool, CP—ceruloplasmin, HEPH—hephaestin, NTBI—non-transferrin-bound iron, FT—ferritin, FT-H—H-ferritin, TIM-1—T-cell immunoglobulin and mucin domain.
The Tf/TfR1 pathway is considered to be the major route for iron transport across the luminal membrane of the capillary endothelium [
107]. According to the widely established transcytosis mechanism, Tf binds to TfR at the luminal side of the brain capillaries [
32]. The complex traverses the cell in the endocytosis vesicle, where the acid environment facilitates the release of ferric iron from Tf and its reduction to ferrous iron by endosomal reductase [
110], possibly duodenal cytochrome b (DCYTB) or six-transmembrane epithelial antigen of the prostate-2 (STEAP2) [
111]. The next steps in this pathway are still not completely clear. One possibility is that ferrous iron is transported from the endosome to the cytosol by the DMT1 [
112] and joins the intracellular labile iron pool (LIP) [
113] (). It could be further utilized for metabolic purposes by the endothelial cells, stored in endothelial cell ferritin [
114] or imported into mitochondria via mitoferrins and TfR2 [
115]. It could be also released into the extracellular fluid by action of export protein ferroportin (FPN1) [
108], and reoxidized to Fe
3+ by ferroxidases hephaestin (HEPH) and ceruloplasmin (CP) [
100]. Studies have confirmed that capillary endothelium of the BBB, neurons, and astrocytes, has the ability to express FPN1 and HEPH [
116,
117]. The alternative mechanism that has been proposed is that the endosome containing Tf-TfR1 complex goes all the way to the abluminal side and releases iron between the endothelial cells and astrocyte end-foot processes [
99]. The released ferrous iron is then oxidized to ferric iron by the ferroxidase activity of CP or HEPH expressed on the end-foot processes [
112]. Oxidized iron binds to apo-Tf circulating within the brain [
113] (). The main source of Tf in the brain interstitium is its diffusion from the ventricles and a certain amount is synthesized in oligodendrocytes [
118]. Because of the low concentrations of Tf in the cerebrospinal fluid (CSF), iron saturation of CSF Tf is almost 100%, while serum Tf is saturated by about 30% [
99]. Consequently, under conditions of IO, CSF Tf has much lower buffering capacity [
119], NTBI levels may be quite high [
120], and the vulnerability of neuronal cells to iron toxicity increases [
119].
Iron may also enter the brain through epithelial cells of the choroid plexus, which form the BCSFB [
121]. The choroid plexus consists of fenestrated capillaries so the holo-Tf can cross them and reach the choroidal epithelium [
122]. Further, the iron is released the same way as from the BBB endothelial cells by means of DMT1, FPN1 and ferroxidases [
14]. When iron enters the CSF, there is no diffusional barrier between CSF and interstitial fluid. Iron binds to Tf in CSF and supplies CNS cells expressing TfR1 [
123].
Different cell types in the brain acquire iron by distinct pathways. Neurons express high levels of TfR1. Therefore, Tf is the main source of iron for neurons [
112], although neurons can also uptake NTBI from interstitial fluid [
124]. Unlike them, oligodendrocytes and astrocytes do not express TfR1 and their main source of iron is NTBI [
110]. Namely, ferrous iron in the brain interstitium can also bind to ATP or citrate released from astrocytes and it is transported to oligodendrocytes and astrocytes as NTBI [
99] ().
Oligodendrocytes acquire NTBI via the T-cell immunoglobulin and mucin domain (Tim-1). It is a ferritin receptor exclusively expressed in oligodendrocytes that binds H-ferritin [
125]. Astrocytes express ferri-reductase on their plasma membranes to reduce ferric to ferrous iron and facilitate iron uptake [
126] (). Once iron enters the brain cells, the iron pool is tightly regulated. It has to provide enough iron for cellular functions and prevent the development of oxidative stress [
110]. Ferritin has an important role in iron sequestration and free iron level reduction [
127], whereas neuromelanin captures large amounts of iron in certain neurons for longer-term storage [
128]. Namely, the pigment neuromelanin acts as a scavenger binding redox-active metal ion such as iron. The expression of ferritin varies in different cell types according to their functional requirements for iron. Neurons contain the least, and microglia contain the most amount of cytosolic ferritin [
129] but in the hypoxic conditions ferritin synthesis increases in cortical neurons and decreases in glial cells [
130]. Ferritin degradation by the autophagy-lysosome system [
131] initiates iron release, mainly through FPN1 [
132]. Since hepcidin regulates the expression of FPN1, it modulates cellular iron level as well [
13]. Recent studies revealed that hepcidin can be produced by the brain endothelium [
108] or systemically derived by passing the BBB [
133], and it is widely distributed in the brain [
134,
135]. Hence, hepcidin may be involved in the regulation of iron availability and circulation in the brain [
108]. Cellular iron levels are also modulated at the post-transcriptional level by binding to the iron response elements (IREs) of mRNA of (iron response proteins (IRPs) [
13].
When some of these cellular and molecular mechanisms of iron regulation are disrupted, the brain iron homeostasis is disturbed as well. If there is either too much or too little iron in the brain, numerous neurologic disorders can occur [
14]. Excessive brain iron accumulation is found in MS, PD and AD, amyotrophic lateral sclerosis (ALS), neurodegeneration with brain iron accumulation, and Huntington’s disease [
114]. ID is associated with significant cognitive, performance and brain structural deficits [
136].
3. Conclusions
Older age is associated with increased risk of ID, elevated body iron stores and increased brain iron levels [
23]. Inadequate iron supply, which often accompanies aging, leads to cerebral hypoxia [
138], insufficient neurotransmitter synthesis [
13], impaired myelination [
139], and consequently to poorer cognition, cognitive decline, and dementia [
141]. On the other hand, brain iron accumulation is considered as a hallmark of aging [
146] and it is associated with the progressive imbalance between antioxidant defenses and intracellular generation of ROS [
147]. This increases susceptibility of aged brain to diseases and thus makes aging a major risk factor for neurodegenerative diseases development [
14]. Therefore, it would be very important for the future research to determine the exact cellular and molecular mechanisms related to perturbations in iron metabolism in the aging brain to distinguish between physiological and pathological aging and find possible therapeutic targets for neurodegenerative diseases.
To counteract the ID during aging, one should certainly consider iron supplements recommended by a physician to correct the anemic state.
However, it should be noted how this supplementation may not be warranted for healthy elderly people consuming a balanced diet. In contrary, it could be detrimental for those who are homozygous or heterozygous for the HFE mutations, since recent studies showed that even moderate increases in body iron may increase the risk for body disorders including neurological ones [
270], or cause irreparable damage to the brain neurons [
137,
271]. Because of that, older people should be careful consuming a high-iron content diet as well.
The major unknown is still the sex-related differences in iron metabolism that come with aging. Increasing experimental and clinical evidence support the idea that neurological disorders differ between women and men, suggesting the existence of different underlying mechanisms involved in their pathogenesis [
16,
17,
18,
19,
20,
21,
22]. However, we are still far away from the actual understanding of what underlies these differences. We need a better understanding of the underlying mechanisms of how sex hormones can influence the iron metabolism and further, the development of neurological disorders. New insights into aging processes, which include the impact of sex hormones on iron metabolism as well, could enlighten the understanding of these differences during aging.