Zika virus (ZIKV) is an arbovirus first discovered in the Americas. ZIKV infection is insidious based on its mild clinical symptoms observed after infection. Currently, no specific prophylactics or therapeutics are clinically available to treat ZIKV infection. Development of a safe and effective vaccine is essential to prevent the rise of any potential pandemic.
1. Introduction
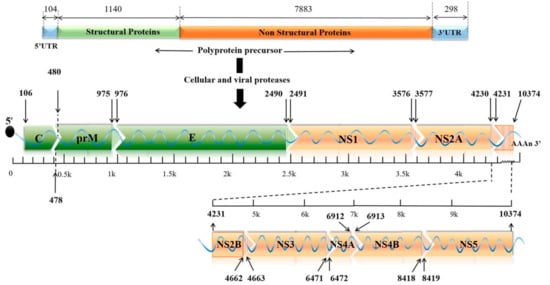
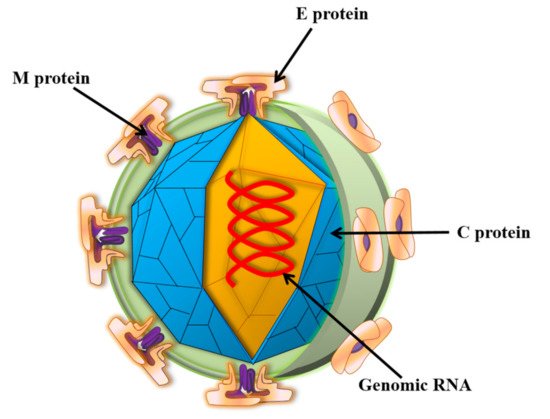
Zika virus (ZIKV) is a small envelope, positive-strand RNA virus belonging to the Flavivirus family of Flaviviridae [1]. As shown in Figure 1, the genome-encoded polyprotein can be cleaved into three structural proteins (capsid (C), anterior membrane (prM), and envelope (E)) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) [2]. The mature ZIKV particles consist of 90 E homodimers and 90 M homodimers on the lipid membrane, and the genomic RNA is surrounded by C protein (Figure 2). E proteins are responsible for receptor binding, attachment, viral entry, and membrane fusion. Transmission routes are arthropod vectors (e.g., Aedes aegypti), intrauterine (perinatal), and through sex and blood-related pathways [3]. ZIKV was also found to be present in the breast milk [4,5]. When ZIKV attacks pregnant women, it is easily reproduced in the placental tissue and seriously affects the fetal central nervous system and immune system, causing congenital Zika syndrome (CZS) in infants [6,7,8]. Studies have identified the major epitopes present on ZIKV structural proteins that can induce neutralizing antibodies [9]. Owing to high structural homology, the presence of common epitopes between Dengue virus (DENV) and ZIKV were reported earlier [10]. However, it is not feasible to prepare a ZIKV vaccine based on the principle of “cross-reactivity of the neutralizing antibodies” [11]. Several in vitro experiments demonstrated an antibody-dependent enhancement (ADE) of ZIKV infection after DENV infection, which poses a challenge to the development of a safe vaccine.
Figure 1. The genomic structure of ZIKV. The genome contains a single ORF encoding three structural proteins (C, prM, and E) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) with two UTRs at both ends. ORF: opening reading frame; C: capsid; prM: premembrane; E: envelope; UTR: untranslated region.
Figure 2. Schematic diagram of ZIKV structure. Zika virions are enveloped, spherical, and approximately 50 nm in diameter. The surface prM/M and E proteins are symmetrically arranged on the viral surface. M: membrane; E: envelope; C: capsid.
2. Development of Vaccines against ZIKV Infection
Development of a safe and an effective vaccine plays an important role in preventing the potential spread and serious harm caused by of ZIKV infection.
2.1. DNA Vaccines against ZIKV Infection
The DNA vaccine platform has been used for over twenty-five years to develop candidate vaccines against numerous pathogens. DNA vaccines can induce both humoral and cellular immune responses and are capable of mediating long-term protection [12]. Most currently developed DNA vaccines for ZIKV contain prM and E genes coding for prM and E proteins (Table 1). Type I interferon receptor alpha-chain null mice (Ifnar1−/− mouse model) [13,14] exposed to ZIKV developed severe damage to the testes and sperm [15,16], but a DNA vaccine encoding ZIKV prM-E completely protected mice against such ZIKV-associated damage [17]. The immunogenicity of a DNA-based vaccine candidate, pVAX1-ZME, expressing the prM/E protein of ZIKV, was evaluated in maternal and post-natal protection of suckling BALB/c mice, and it was demonstrated that the administration of three doses with 50 µg of pVAX1-ZME by in vivo electroporation induced robust ZIKV-specific cellular and long-term humoral immune responses with high and sustained neutralizing activity in adult BALB/c mice. The neutralizing antibodies passively protected against ZIKV infection in neonatal mice and effectively inhibited delay in growth [18]. GLS-5700, a DNA-based vaccine that encodes the prM and E antigenic regions of ZIKV, was shown to prevent fertility loss in male IFNAR−/− mice [19]. The Vaccine Research Center (VRC) of the National Institute of Allergy and Infectious Diseases (NIAID) and National Institutes of Health (NIH) in USA have developed two DNA vaccine candidates, named VRC5288 and VRC5283, and tested them in phase I clinical trials to assess their safety, tolerability, and immunogenicity in humans [20]. Another DNA vaccine was developed by using a single tetrafunctional amphiphilic block copolymer (ABC) encoding the full sequence of prM-E, which induced a high neutralizing antibody titer against three divergent ZIKV isolates in six-week-old female C57BL/6C mice [21].
DNA vaccines offer optimization of the sequences of the encoding genes in a flexible manner and the ability to test multiple candidate antigens rapidly [
23]. They are chemically stable and easy to produce, with no requirement for cold-chain storage, and they are also cost-effective to manufacture on a large scale [
22,
24]. However, DNA vaccines involve expression of multiple foreign genes and thus have the potential of integrating the exogenous gene into the host genome, leading to induction of host autoimmunity [
25].
2.2. Subunit Vaccines against ZIKV Infection
Based on their rapid, stable, and consistent production capabilities, subunit vaccines are considered as effective tools to prevent virus infection. Subunit vaccines against ZIKV have been designed and tested in animal models (Table 2). Viral structural proteins, such as E protein and its domain III (EDIII), are attractive vaccine targets. The immunogenicity of a subunit vaccine candidate comprised of ZIKV E protein and two clinical adjuvants (Alum and CoVaccine HTTM) was evaluated in Swiss Webster, BALB/c, and C57BL/6 mice [26]. These vaccine formulations induced robust IgG titers and high levels of neutralizing antibodies in all three mouse strains and protected them against viremia after ZIKV infection [26]. Another research team used a recombinant subunit platform consisting of antigens produced in Drosophila melanogaster S2 cells to develop two candidate formulations. The first formulation contained 25 µg of ZIKV E which was adjuvanted with 10 mg Co-Vaccine HTTM, and the second formulation contained 50 µg of ZIKV E protein with Alhydrogel® 85 plus 1.2 mg of elemental aluminum. High neutralizing antibody titers were induced in a non-human primate (NHP) viremia model, and passive transfer of the plasma from the macaques protected against viremia in ZIKV-infected BALB/c mice [27]. Based on this observation, another research team agreed that high anti-ZIKV titers protected against viremia, but they also suggested that low titers could provide an incremental degree of protection, albeit not sufficient to prevent viremia [26]. Purified EDIII from transformed Escherichia coli inclusions induced high titers of IgG and ZIKV neutralizing antibodies, which showed no evidence of ADE induction in C57BL/6 mice [28].
A truncated subunit vaccine consisting of the first 450 amino acids at the N-terminal region of the ZIKV FSS13025 strain E protein (E90) was investigated in 7- to 8-week-old CD-1 (ICR) immunocompetent mice for both in utero and neonatal ZIKV infection. Results demonstrated that immunization of pregnant mice with E90 protected the developing brains of offspring, both in utero and during the neonatal period, from subsequent ZIKV infection and microcephaly. Most importantly, E90 vaccination protected mice from ZIKV infection, even at 140 days post-immunization [29]. Another group showed robust induction of ZIKV-specific humoral response in adult BALB/c mice by E90, and passive transfer of the antisera from these mice conferred absolute protection against lethal ZIKV challenge in a neonatal mouse model [30]. These studies demonstrated the promising nature of recombinant ZIKV E90 as a ZIKV subunit vaccine that deserves further clinical development. One study used seven-day-old male and female BALB/c pups, 6 to 8-week-old female BALB/c mice, and 5-week-old male and female type-I IFN receptor–deficient A129 mice to investigate the long-term immunogenicity and neutralizing activity of the ZIKV EDIII fragments. The results showed that ZIKV EDIII fragment, especially E298–409, could induce sustained development of neutralizing antibodies [31]. The E298–409-specific antibodies upon passive transfer prevented ZIKV infection in newborns and immunocompromised adults [31]. Thus, this subunit vaccine based on the critical fragment (E298–409) of ZIKV EDIII is one of the promising vaccine candidates for ZIKV infection. It has been proven that the recombinant ZIKV subunit vaccine is a safe and efficacious option for the prevention of ZIKV infection. However, its less immunogenic nature is the major disadvantage, requiring more doses and appropriate adjuvants [32].
2.3. Live-Attenuated Vaccines against ZIKV Infection
Live-attenuated vaccines (LAV) are those that reduce the infectivity of pathogens after various treatments, but retain their immunogenicity. A number of live-attenuated vaccines have been evaluated in animal models (
Table 3). Candidate LAV viruses with engineered deletions in the 3′ untranslated region (UTR) provide immunity and protection in animal models of ZIKV infection [
33]. Recently, a LAV candidate containing a 10-nucleotide deletion in the 3′ UTR of the ZIKV genome (10-del ZIKV) was developed. Even when immunized at a low dose with only 10-del ZIKV, complete protection from viremia by the induction of a high level of neutralizing antibodies was observed, preventing a decrease in the sperm count in A129 mice [
34]. Another LAV candidate containing a 20-nucleotide deletion in the 3′ UTR of the ZIKV genome prevented viral transmission during pregnancy and testis damage in mice, as well as infection of NHP [
35]. The ZIKV-3′UTR-Δ20-LAV is less sensitive to type-I-interferon inhibition than ZIKV-3′UTR-Δ10-LAV [
34], making it more virulent than ZIKV-3′ UTR-Δ10-LAV in A129 mice. Collectively, LAV candidates containing deletions in the 3′ UTR of the ZIKV genome are efficacious and have the potential to be promising vaccine candidates.
A live-attenuated ZIKV vaccine candidate encoding NS1 protein, but without glycosylation (ZIKV-NS1-LAV), was demonstrated to markedly diminish viral RNA levels in maternal, placental, and fetal tissues, which resulted in protection against placental damage and fetal death [
36]. A single-dose LAV candidate containing a 9-amino-acid deletion in the viral capsid protein that infects cells with controlled, limited infection rounds was developed to test its safety and immunogenicity in A129 mice [
37]. The results showed that a single-dose immunization of this LAV vaccine elicited protective immunity that completely prevented viremia, morbidity, and mortality. At the same time, it also fully prevented infection of pregnant mice and maternal-to-fetal transmission. Interestingly, injection of this vaccine with 10
4 plaque-forming units to 1-day-old mice did not cause any disease or death, underscoring its safety [
37].
2.4. Virus-Vector-Based Vaccines against ZIKV Infection
Virus-vector-based vaccines of ZIKV are designed to introduce ZIKV genetic material into cells using a virus as the carrier to induce protective immunity and achieve lasting protection (
Table 4). Lentivirus, retrovirus, and adeno-associated virus can be used as carriers. Adenovirus vectors (AAV) can reduce the risk of insertion mutagenesis, induce strong innate immune and adaptive immune responses in mammalian hosts, and are easy to use for genetic modifications [
38]. Thus, AAVs are widely used in the treatment of infectious diseases. Adenovirus-vectored vaccines represent a favorable controlling strategy for the ZIKV epidemic. Two adenovirus-vectored Zika vaccines were constructed by inserting a ZIKV prM-E gene expression cassette into human adenovirus type 4 (Ad4-prM-E) and 5 (Ad5-prM-E) vectors [
39]. A replication-defective vector which contains the full length of prM-E genes of ZIKV PRVABC59 strain [
39] was created. Surprisingly, ELISA and plaque reduction neutralization tests showed negligible levels of anti-ZIKV antibodies after Ad4-prM-E-vaccination in C57BL/6 mice, suggesting that Ad4-prM-E vaccination induces only T-cell responses, whereas Ad5-prM-E vaccination induced both anti-ZIKV antibody and T-cell response [
39]. Interestingly, coadministration of UV-inactivated Ad4 vector with Ad5-prM-E vaccine led to significant reduction in CTL and overall T-cell responses, compared to Ad5-prM-E alone [
40], highlighting the differences in serotype-specific immunity induced by adenovirus vectors.
2.5. Purified Inactivated Zika Vaccines (PIZV)
The inactivated virus vaccine is produced by killing the original live virus through heat or chemicals and then introducing the remaining virus shell into the host body. Different inactivation strategies are used, such as UV, formalin and iodonaphthyl azide. It was reported that 50 and 100 µM of iodonaphthyl azide could completely inactivate ZIKV [
58]. A number of PIZV candidates have been evaluated in animal models (
Table 5).
Two-dose immunization of alum/adjuvant-added, purified, inactivated ZIKV vaccine (PIZV) protected AG129 mice against lethal ZIKV challenge [
59]. In addition, passive immunization of naïve mice with anti-ZIKV-immune serum showed a strong positive correlation between neutralization antibody titers and protection against lethal challenge with ZIKV [
59]. PIZV with aluminum hydroxide, developed by the Walter Reed Army Institute of Research and further optimized by Sanofi Pasteur, induced robust neutralizing antibody responses and provided absolute protection from challenge with a homologous ZIKV strain in immunocompetent BALB/c mice and in Cynomolgus macaques [
60,
61]. Two-dose vaccination of PIZV at varying concentrations, ranging from 0.016 µg to 10 µg, elicited a dose-dependent and long-lasting neutralizing antibody response in Indian rhesus macaques [
62]. Complete protection against ZIKV infection was achieved with the higher PIZV doses of 0.4 µg, 2 µg, and 10 µg at 6 weeks and with 10 µg PIZV at 1-year post-vaccination [
62]. Two doses of PIZV gave robust protection against ZIKV challenge in rhesus monkeys at 1-year post-vaccination [
63].
2.6. Virus-Like Particle (VLP)-Based Vaccines against ZIKV Infection
Virus-like particles (VLPs) are noninfectious because they are empty shell structures having no viral genome. Many viral structural proteins have the ability to assemble automatically into VLPs. They can be produced in a variety of expression systems, such as suspension cultures of mammalian, yeast and insect cells. A number of VLP-based ZIKV vaccines have been tested in animal models (Table 6).
VLPs were successfully produced by co-expression of the ZIKV structural proteins C-prM-E together with a truncated form of the protease NS3Pro linked to its cofactor NS2B constituting the viral NS2B/NS3Pro protease complex [
67]. Recent studies have demonstrated that co-expression of ZIKV C-prM-E and ZIKV NS2B/NS3 [
67] or WNV NS2B/NS3 protease [
68] has facilitated the cleavage of C and prM, thereby allowing more efficient production of VLPs. Negative staining studies revealed that both VLPs and real virus particles are similar in size, morphology, as well as surface appearance. Indeed, the major surface glycoprotein E of ZIKV is present on the VLP surface [
67], and E protein is the major target for neutralizing antibodies [
68].
VLPs mimic the conformation of natural viruses by expressing one or more structural proteins and thus stimulate robust antibodies in vivo. Antigens are present in their native conformation, but VLPs, to their advantage, use no replication virus [
32]. However, application of VLP-based vaccine candidates for clinical use needs further studies.
2.7. mRNA-Based Vaccines against ZIKV Infection
Synthetic messenger RNAs (mRNAs) have emerged as a versatile and highly effective vaccine platform for encoding viral antigens, and they are quite attractive because of their production rapidity and flexibility. Some mRNA-based vaccines against ZIKV infection have been developed and tested in animal models (Table 6).
2.8. Other Types of Vaccines against ZIKV Infection
Apart from the above-mentioned vaccine platforms, other types of vaccine candidates are also available against ZIKV. A chimeric ZIKV with DENV-2 prM-E genes was highly potent in A129 mice, and it induced robust neutralizing antibody responses, which conferred complete protection from challenge with ZIKV [
75]. Over the last several years, computer-assisted peptide vaccines have started to draw much attention as alternative vaccine candidates. The peptide vaccine is a more focused approach to precisely locate the epitope region within the antigens and elicit immune responses. Occasionally, multiple peptides for one or more viral infections can be combined into clusters for immunization against a wide spectrum of infections [
76].
The potential advantages and disadvantages of the above mentioned different types of vaccines are summarized in Table 7.
3. Conclusions
Currently, more efforts are needed to prevent and/or treat ZIKV infection. The development of a safe and efficacious ZIKV vaccine remains a global health priority. Vaccine development and use on a large scale to prevent pandemics involve several factors such as mass production of vaccine, adjuvant selection, establishment of optimal animal models for preclinical studies, validation of safety and efficacy in animal models and clinical trials in different parts of the world using a large cohort of patients, immunization strategies, storage conditions, as well as manufacturing and production costs. Novel proposals for multi-epitope vaccines, as well as the discovery of new adjuvant formulations and delivery systems that could enhance and/or modulate immune responses, may help pave the way for development of successful vaccine candidates.
This entry is adapted from the peer-reviewed paper 10.3390/vaccines9091004