1. Introduction
Population aging is now a global phenomenon that is rapidly evolving all over the world. In the European Union, the number of people over sixty-five is expected to increase from 85 million in 2008 to 151 million in 2060 [
1]. Worldwide, people aged over 65 are estimated to increase from 461 million in 2004 to 2 billion by 2050 [
2,
3]. This will have important consequences for the planning and delivery of health care and social assistance services. A greater life expectancy, in fact, leads us to reconsider not only the condition of the elderly, but also what kind of implications aging will have in our lifetime [
4,
5]. That is, the ability to dedicate ourselves to our activities will be no different from that of a younger person, wherever we spend our old age in good health. On the contrary, where the years of old age are dominated by loss of health, cognitive decline and therefore, the loss of self-sufficiency, the implications for individuals and for society as a whole would be much more negative [
5]. Especially in high-income countries, it is now customary for many people to spend their old age in innovative ways, such as starting a new career or even starting new studies, or devoting oneself completely to a neglected passion during working age [
6]. It is clear, however, that the extent of these new opportunities arising from these long years of life depends, for everyone, on maintaining health and self-sufficiency.
Healthy behaviors in middle age predispose to so-called “healthy and successful” aging, which is the condition of the absence of serious chronic diseases or of important decline in cognitive or physical functions, or mental health [
7]. Healthy behaviors include following a healthy diet, with an adequate caloric restriction to the state of health and physical activity, not smoking, intake of moderate amounts of alcohol, especially in women. Akbaraly et al. [
8], and Samieri et al. [
9] from the results of the Whitehall II cohort study and of the Nurses’ Health Study, respectively, showed that a healthy dietary pattern, like the Mediterranean Diet (MD) pattern, was associated to a lower incidence of chronic disease and to a lower physical impairment in old age. In addition, they showed that a healthy dietary pattern was associated to a lower cardiovascular risk and to a lower risk of premature death.
2. Aging and Frailty
The frailty of the elderly is a condition characterized by an increased vulnerability to poor homeostasis resolution after a stress event, which increases the risk of negative outcomes, including falls, delirium and disability. Most geriatricians intuitively recognize frailty; otherwise, this is mostly misunderstood, or confused with the presence of comorbidity and disability [
10,
11,
12]. With the aim to provide a standardized definition for frailty, Fried et al. [
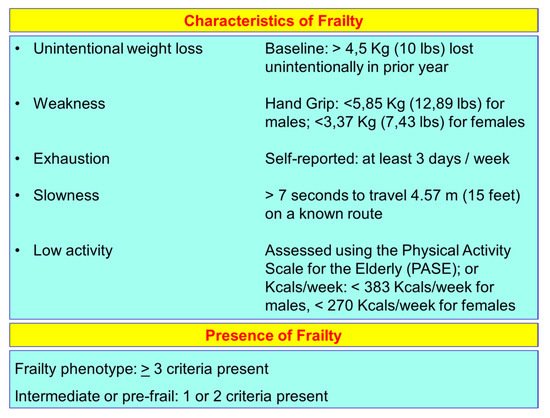
13] identified the “frailty phenotype”, which is the measure most frequently used. Fried’s criteria of frailty considers five items to determine the level of frailty: weight loss, exhaustion, low physical activity, slowness, and weakness (
Figure 1). Rockwood et al. [
14] starting from the identification, through the Canadian Study on Health and Aging (CSHA), of 70 items including signs, symptoms and abnormal tests that characterized the fragility, built their “CSHA Clinical Frailty Scale”, which considered seven levels, form Very fit to Severely frail (
Figure 2). More correctly, frailty develops due to the greater decline, already linked to age, up to the severe impairment and the appearance of pathological states, of the different organs and systems, which overall, leads to a condition of greater vulnerability and reduced resilience and ultimately, to sudden and severe health changes triggered by even mild stressors. In the frail elderly an apparently small insult, for example, a small infection or minor surgery produces a significant worsening of the patient’s condition. This means that the frail elderly loses all self-sufficiency, undergoes a hypokinetic syndrome to the condition of entrapment, increases the risk of falls, and develops a state of confusion, with a serious deterioration of cognitive functions. [
15]. Frail people have several functional deficits, which often cause falls, immobility and confusion. They are the patients who come to the hospital with greater frequency and are the main recipients of long-term care services. [
16]. About 25–50% of people over the age of 85 are estimated to have different degrees of frailty; these are subjects at high risk of falling, permanent disability and death, in most cases requiring long-term care [
13,
15]. However, if we consider that 25–50% of people over the age of 85 are frail, we must consider that that 50% to 75% of people over 85 years are not frail, which raises several questions, namely: How does frailty develop? How we can prevent it? How can we reliably detect it?
Figure 1. Fried’s criteria of frailty (line 69).
Figure 2. The Canadian Study of Health and Aging (CSHA) Clinical Frailty Scale by Rockwood.
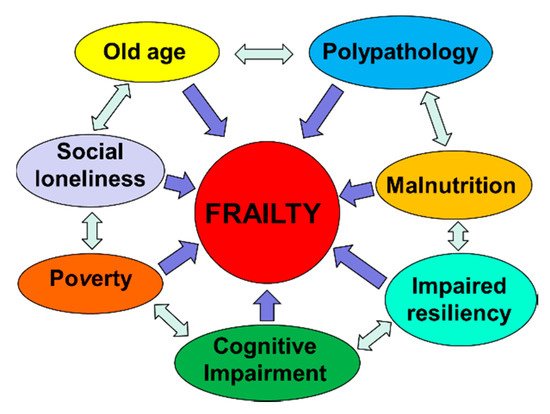
Frailty must not be understood as an inevitable consequence of aging, but rather, must be understood as a geriatric syndrome [
13,
17,
18,
19]. It is therefore more appropriate to speak of “frailty syndrome”, i.e., a chronic pathological condition that results from the interaction of various factors, including aging-related physiological alterations, poly-pathology, nutritional deficiencies up to severe malnutrition, and the negative impact of socio-environmental factors (
Figure 3).
Figure 3. The vicious circle of frailty (line 92).
3. Aging, or Cellular Senescence, and Health
If we consider aging solely from the biological point of view, we refer essentially to the physiological and progressive accumulation of senescent cells in tissues and organs, which occurs during the life of each individual [
19], with the progressive slowing down and, in some cases, loss of function. The antagonist pleiotropic genes are a set of genes that regulate cellular senescence [
20,
21,
22,
23]; these genes play an important role in the prevention of malignant cell degeneration in the pre-neoplastic degeneration, i.e., eliminating cells from the cell cycle; the same genes are also involved in the mechanisms of protection in the physiological cellular senescence processes and towards age-related diseases [
24,
25,
26].
The reduction of the capacity of cell regeneration and tissue repair are the basis of the physiological senescence process, substantially due to the slowdown process, up to the complete stop of the cycle of growth and replication of the progenitor cells. These aging cells also produce a series of proinflammatory and lytic molecules of the extracellular matrix in the process known as the secretory phenotype associated to senescence (SASP). Consequently, those mechanisms normally needed to maintain tissue homeostasis in the aging organism produce a series of alterations in the structure and function of the cells, resulting in degeneration and pathological senescence. The aging process also involves the immune system; in particular, the cell-mediated defense mechanism is slowed down. Furthermore, senescent cells do not produce enough signals to activate immune cells. As a result, a few aging senescent cells survive in the body. The senescent cells, in addition to reducing the functional efficiency of the organs and systems during aging, make them more vulnerable, therefore subject to further deterioration, after exposure to environmental stress factors [
27].
In physiological aging, senescence is induced by the accumulation of different degenerative factors that slowly accumulate in the cells and that are responsible for macromolecular damage [
28,
29,
30], for example, the shortening of telomeres, the addition of secondary DNA alterations to oxidative damage, stress degeneration of the endoplasmic reticulum (ER). The disease takes place when environmental stressors attack the tissues already in the presence of senescent cells with very low resilience capacities [
31,
32,
33]. The stress that can cause the disease can be foreign and abnormal to the cell, as damaging DNA agents in cigarette smoke; otherwise, it can be a more intense response to a stressor event prolonged over time by the same factors that work in physiological aging, e.g., erosion of telomeres in lung epithelial cells chronically damaged by smoke [
34]. It is therefore evident that aging is the result of a multi-factorial interaction between local and systemic environmental factors, and involutional factors due to cell senescence. Over time, these environmental stressors add to the cellular changes linked to physiological aging, decreasing resistance to stress and further reducing tissue resilience. That is, the chronic stimulation of a stressor factor on a tissue already in a condition of cellular senescence is more likely to induce the onset of a pathological condition [
27].
4. The Role of Senescence in the Progression of Diabetes Mellitus and Atherosclerosis
Under physiological conditions, the synthesis and release of insulin by pancreatic beta cells, based on glycemic levels, allows the absorption of glucose in sensitive peripheral tissues, i.e., the liver and skeletal muscle, with the consequent production of energy by means of aerobic glycolysis [
35]. In obese people, build-up of abdominal fat tissue, which is characterized by a chronic inflammatory state, and the excess of free circulating fatty acids, can cause a state of peripheral tissue insulin resistance [
36,
37]. Thus, what occurs is a condition of increased and abnormal prolonged demand in the time of production and release of insulin with the consequent prolonged stimulation of pancreatic beta cells. The increased replication activity, and consequently, the wear of telomeres, leads to premature senescence of beta cells [
38], thus reducing the efficacy of the reaction to hyperglycaemia with the consequent aggravation of type 2 diabetes mellitus (T2DM) and the onset of insulin resistance. This reduced ability to regulate blood glucose levels alters the metabolic processes of the cells and speeds up the aging process. This glucose-induced cellular toxicity induces a condition of chronic stress and consequently, pathological senescence in various cell lines, for example, fibroblasts, cells of the renal tubules, endothelial cells, mesenchymal cells [
39,
40,
41,
42], and even neurons. Therefore, we see the appearance and progression of a number of diseases, such as atherosclerosis, chronic renal failure [
43] and Alzheimer’s disease. Furthermore, the accumulation of abdominal adipose tissue and the release in circulation of excess free fatty acids can also cause the progression of atherosclerosis. During the formation and progression of plaque, the proliferation of smooth muscles and the reduction of endothelial nitric oxide synthase levels can lead to the shortening of telomeres and increased oxidative stress, respectively [
44,
45]. Plaque progression is therefore the result of a series of interactions between smooth muscle cells and immune system cells, modulated by the synthesis and release of cytokines and adhesion molecules by endothelial cells, smooth muscle cells and immune cells. The complexity of the action mechanisms of senescent cells in the atherogenesis process is evident. Paradoxically, senescence in atherosclerosis acts as an initial protective factor by restricting and slowing the development of plaques and minimizing plaque-disrupting apoptosis. After a limit threshold of senescence burden, the proinflammatory matrix-degrading SASP exacerbates disease [
27].
5. Caloric Restriction, Effects on Metabolism of Adipose Tissue and Increase of Longevity
It is known that overweight at young and middle age, by decreasing insulin sensitivity, can accelerate the aging process and predispose to the onset of age-related diseases [
45,
46,
47,
48]. In experimental models conducted both in mice and in humans, it has been observed that caloric restriction (CR) produces significant effects in terms of weight reduction; much of the weight loss comes from the reduction of white adipose tissue (WAT) and visceral fat deposits [
45,
49,
50]. It is true that the common strains of laboratory rodents all have an extended lifespan in response to CR, however, this is not a universal effect. Interestingly, some inbred mice strains show only a modest effect on extension of the life span after CR. In some cases, a reduction in life span has even been observed [
51]. We can explain these apparently conflicting results considering that the effect of CR on the metabolism is a decrease in carbohydrate dependence and a greater use of fatty acid oxidation to produce energy [
52]. Indeed, the level of fat reduction following dietary restriction remains a key component of the beneficial effects of CR. In a study conducted on 40 strains of mice, increased longevity induced by CR was more evident in old mice (15–17 months of age) that, with a 40% CR diet, were able to retain more fat deposits, showing that retainment of both fat mass and lean mass in aging can correlate with more longevity [
53]. Subsequently, the results of a study conducted on progeroid mice showed that a caloric restriction regimen of 30% increased its median duration and the maximum residual life expectancy by three times. Specifically, mice subjected to caloric restriction retained 50% more neurons and maintained full motor function, far beyond the lifespan of ad libitum-fed mice.
6. Caloric Restriction and Inflammatory State
Gene expression analysis has shown that dietary restriction increases DNA resistance to stress-induced damage, as well as improving antioxidant defenses and metabolic processes primarily mediated by insulin, as well as by other hormonal signaling pathways. Dietary restriction also affects mitochondria activity, modulates apoptotic response and modifies pro-inflammatory and anti-inflammatory cytokine production [
54]. The results of further randomized human studies confirm that caloric restriction stimulates the body to adapt to the use of any available energy substrate, both glucose and fatty acids [
55]. The term “metabolic flexibility” optimally defines this adaptation property, which has long been linked to metabolic health and longevity [
56]. Studies conducted on primates have also shown that a strong relationship exists between CR and the reduction of the pro-inflammatory state [
57]. A subsequent study linked the reduction of the pro-inflammatory state to the inhibition of the inflammasome, induced, in turn, by the increase in serum levels of β-hydroxybutyrate during fasting [
58]. Another suggested a mechanism through which CR could suppress the harmful activation of the immune system, which concerns the preservation of the thyme function induced by the CR, thus preserving the function of T cells [
59], or a reduction of inflammation induced by oxidative stress [
60].
7. Caloric Restriction, Mitochondria Activity and Reactive Oxygen Species Production
Numerous experimental models conducted on animal organisms have involved mitochondrial proteins encoded by the nucleus in the life span adjustment process [
61,
62]. These experiments on genes mostly involved an alteration of the electron transport chain process with consequent impairment of mitochondrial function; nevertheless, they have led to a greater life expectancy [
48]. Conversely, in mice models bearing significant mitochondrial genome mutations designed to significantly impair mitochondrial function, a phenotype characterized by accelerated aging was observed [
63,
64]. The seemingly contradictory results observed in these experimental models may be due to the variability of the alteration of mitochondrial function that was obtained from time to time. Experiments conducted on C. elegans have shown an increase in life expectancy as a result of a modest reduction of the various components of the mitochondrial electron transport, while a significant reduction, i.e., a sign of high mitochondrial damage, was associated with a reduction in the lifespan [
65]. Evidence from mammalian models has shown that a slight reduction in mitochondrial function is associated with prolonging lifespan. This assumption seems to contradict the hypothesis that reactive oxygen species (ROS) support aging [
66]. In fact, although mitochondrial ROS can be dangerous for life [
67], it is equally clear that a slight stimulation of mitochondrial ROS production can stimulate the activation of the anti-redox mechanism with protective effects in the long term.
8. Caloric Restriction, Hormesis and Mitochondria Activity during Aging
“Hormesis”, from the Greek verb “ormao” that means “to stimulate”, is described as the adaptive function of a cell or an organism characterized by a biphasic dose-dependent response, following exposure to a variety of stimuli, such as toxins or other stressors. The hormetic responses, generally, show a modest stimulation of the response of the organism to low doses of a stressor, or an inhibition of the same response to high doses [
68]. “Mitochesis” is defined as the hermetic response of the mitochondria, i.e., that the activation of a low level of stress can protect against major and subsequent stress [
69,
70,
71]. It is therefore clear that ROS levels are not the only factor responsible for mitochondrial stress affecting life span. Another mechanism linking the aging of mitochondria is constituted by the turnover capacity, that is, the balance between biogenesis, or the synthesis of new mitochondria, and mitophagy, or the removal of aged and damaged mitochondria. It is now clear that all interventions, such as CR, which result in greater longevity, act by inducing mitochondrial biogenesis through the expression of peroxisome proliferation-activated receptor gamma (PPAR-γ), and coactivator 1 alpha (PGC-1α) [
72,
73]. It is equally clear that the reduction of biogenesis contributes to the onset of age-related diseases, both through the reduced activity of PGC-1αand through other routes. [
74]. A gradual reduction in autophagic activity associated with aging was also observed [
75]; a similar age-related decline is also likely to affect the more specialized process of mitophagy. On the contrary, in experimental models, the increase in the expression of gene products that stimulate mitochondrial activity has been observed to determine a prolongation of life expectancy [
76].
9. Caloric Restriction and DNA Methylation
In addition, DNA methylation (methylation drift) is an important epigenetic modification that involves the addition of a methyl group in the cytosine 5 position by DNA methyltransferase to form 5-methylcyctosine (5-mC). This epigenetic mark has the power to activate or deactivate genes and can be inherited through cell division. DNA methylation plays an important role in normal human development, aging, tumorigenesis and other genetic and epigenetic diseases [
48,
77,
78]. Maegawa et al. [
79] found that DNA methylation drifts with age in both mice and humans and monkeys. They also found that Methylation drift is inversely proportional to lifespan across these three-mammalian species. In their elegant study, authors also found that a sample of 22 to 30-year-old rhesus monkeys exposed to 30% CR since 7–14 years of age showed attenuation of age-related methylation drift compared to ad libitum-fed controls, such that their blood methylation age appeared 7 years younger than their chronologic age. They observed the same and more pronounced effects in 2.7–3.2-year-old mice exposed to 40% CR starting at 0.3 years of age.
10. Caloric Restriction, Metabolic Adaptation and Oxidative Damage
Readman et al., in the CALERIE Study [
80]., delivered a highly controlled and intensive behavioral trial targeting a 25% CR diet over 2 years in a sample of 53 voluntary and non-obese adults (34 CR and 19 controls); then, they measured, in a room calorimeter, the component of daily sedentary energy expenditure, i.e., the energy metabolism during sleep. They also measured hormonal mediators of metabolism, including leptin and thyroid hormones, along with urinary F2-isoprostane excretion as an index of oxidative damage. After two years, subjects in the CR group experienced an average weight loss of 8.7 Kg, whereas subjects in the control group maintained weight during the 2-year period. In the CR group, the greater weight loss was from fat mass after two years (−3.2 Kg; p > 0.001). In addition, the authors found a significative reduction of 24-h Energy Expenditure (−186 Kcal/day; p < 0.05), and a significative reduction in Sleep Energy Expenditure (−160 Kcal/day; p < 0.05) in the CR group after two years, indicating a metabolic adaptation to the CR. After adjustment for changes in body composition, while Sleep Energy Expenditure was significantly decreased in the CR group after two years (p < 0.05), the 24-h Energy Expenditure did not decrease significantly after two years, compared to the control group (p > 0.55). As regards the biomarkers of Energy Metabolism and aging, authors found a significant reduction in the CR group after two years of T3 (−0.73 ng/dl; p < 0.05), T4 (−0.16 mcg/dl; p < 0.05), and leptin (−9.3 ng/dl; p < 0.05); in addition, they observed a reduction of fasting insulin and of nigh time core body temperature. Regarding the markers of oxidative stress, the authors found a significative reduction of the urinary excretion of four F2-isoprostane isomers after two years in the CR group (−0.49 ng/mg CR; p < 0.05). The authors have linked these data to the metabolic adaptation of the 24-h Energy Expenditure after the CR. The author also linked the CR with an improved mitochondrial function, with a decreased total body oxygen consumption, and therefore, with a decreased reduction of Oxidative Stress.
11. Mediterranean Diet, Cardiovascular Disease and Mortality
The traditional Mediterranean Diet (MD) is characterized by a high intake of foods of plant origin (fruit, vegetables, breads, other cereals, potatoes, beans, nuts, and seeds) and fresh fruit. Olive oil, namely extra-virgin olive oil, is the main source of fat. Dairy products (mainly light cheeses and yogurt), fish and poultry are consumed in medium-low quantities; particularly, fish is an excellent source of polyunsaturated fatty acids (PUFAs), particularly omega-3 fatty acids; egg consumption is limited to a maximum of four per week; red meat is consumed sporadically and in small quantities, however, no more than once a week. MD has a very low saturated fat content, which represents no more than 8–10% of the total caloric intake. Caloric intake from lipids is not more than 30% of total caloric intake. Wine is usually consumed with meals, but always in moderate doses (1–2 glasses) [
81,
82]. Ancel Keys first demonstrated the health benefits derived from the MD [
83]; thanks to early studies of Ancel Keys, MD has been proposed as a healthy dietary pattern associated with a lower risk of developing cardiovascular and metabolic diseases. The traditional MD has been proposed as a food model to achieve or maintain optimal weight. Trichopoulou et al. [
84] showed that high adherence to the MD, assessed by the Mediterranean Diet Score (MDS), was related to a significative reduction of total mortality [
85].
Subsequently, the PREDIMED study [
86] confirmed the finding above. The authors have shown that all subjects at high cardiovascular risk who followed an MD pattern, supplemented with monounsaturated fatty acids and antioxidants, that is, extra-virgin olive oil (EVOO), or with omega-3 poly-unsaturated fatty acids, that is nuts, had a reduced Hazard Risk (HD) of acute myocardial infarction, stroke or death from any cardiovascular event (MD with EVOO: HR = 0.70, 95% CI: 0.53–0.91, p = 0.009; MD with nuts: HR = 0.70, 95% CI: 0.53–0.94, p = 0.02). They did not observe any effect on reduction of all-cause mortality (MD with EVOO: HR = 0.81, 95% CI: 0.63–1.05, p = 0.11; MD with nuts: HR = 0.95, 95% CI: 0.73–1.23, p = 0.68).
The authors subsequently identified protocol deviations, including the enrolment of household members without randomization, assignment to a study group without randomization of some participants at one of 11 study sites, and apparent inconsistent use of randomization tables at another site. Then, they published a new revised report, which was based on analyses that do not rely exclusively on the assumption that all the participants were randomly assigned [
87]. In their second report, the authors confirmed the lower risk of acute myocardial infarction, ischemic stroke, or death from all cardiovascular event among subjects who were assigned to an MD pattern respect to control subjects who were assigned to a low-fat diet.
A subsequent meta-analysis [
88] analyzed the relationship between adherence to MD and mortality and incidence of each disease; 1,574,299 subjects were involved; they were followed for a time ranging from three to 18 years. The authors demonstrated a significant association between greater adherence to MD, a significant improvement in health status and a significant reduction in mortality Rate Risk (RR) (RR = 0.91, 95% CI: 0.89–0.94; p < 0.0001). The authors have also shown that a greater adherence to an MD pattern was associated with a significant reduction in mortality due to coronary heart disease (CHD) (RR = 0.91, 95% CI: 0.87–0.95, p < 0.0001) and to cancer (RR = 0.94; 95% CI: 0.92–0.96; p < 0.0001).
A further meta-analysis conducted by the same authors [
89] further showed that a great adherence to MD was associated with an improvement in health status and quality of life and to a significant reduction in overall mortality (RR = 0.92, 95% CI: 0.90–0.94, p < 0.00001). Specifically, authors showed a significative reduction of mortality from coronary heart disease (RR = 0.90; 95% CI: 0.87–0.93; p < 0.00001) and from cancer (RR = 0.94; 95% CI: 0.92–0.96; p < 0.00001). A high adherence to MD was also associated with a significant reduction in the incidence of neurodegenerative diseases (RR = 0.87; 95% CI: 0.81–0.94; p < 0.00001), including Alzheimer’s disease.
Nevertheless, as already emphasized in an excellent editorial by Voelker [
90], it would be simplistic to consider the MD as a simple semi-vegan diet rich in fibers, antioxidants and proteins of vegetable origin. We observe the benefits of the MD in its cultural context, where food is part of a lifestyle.
In 2011, the Mediterranean Diet Foundation together with the forum on Mediterranean food cultures developed a consensus document that revised the pyramid of the Mediterranean diet, including cultural and lifestyle elements [
91]. The authors underlined the aspects of socialization, by writing, “The aspect of conviviality is important for the social and cultural value of the meal, beyond nutritional aspects. Cooking, sitting around the table and sharing food in company of family and friends is a social support and gives a sense of community. Make cooking an important activity taking the proper time and space. Cooking can be relaxing, fun and can be done with family, friends and the loved ones’’. At least 30 min of moderate exercise during the day “as a dietary supplement” and adequate rest at night and during the day in the form of a nap after a meal are also recommended in the consent document.
Lastly, Kromhout et al. [
92], further confirmed how a food model according to the features of the MD, assessed through the Mediterranean Adequacy Index, was inversely associated with mortality due to ischemic heart disease (r = −0.91). In particular, the authors confirmed the protective role of cereals (r = −0.52), vegetables (r = −0.52) and legumes (r = −0.62), in addition to the intake of moderate amounts of alcohol in the diet (r = −0.54). The authors also confirmed the association between ischemic mortality of heart disease and dietary intake of high amounts of saturated fatty acids (r = 0.83), whole milk (r = 0.84), confectionery preparations based on simple sugars (r = 0.69), animal meats, with the exception of fish (r = 0.68), and preparations based on animal meats and simple sugars, for example, processed meats (r = 0.84).
Table 1 summarizes the studies we examined.
Table 1. Mediterranean Diet adherence and risk of mortality.
|
Author and Year of Publication
|
Study Design
|
Sample Size
|
Risk of Mortality
|
|
Trichopoulou, 2003, [84]
|
Population-based, prospective study
|
8895 men and 13,148 women
|
Death from any cause:
HR = 0.75 (95% CI: 0.64–0.87) for a Two-Point Increase in the Mediterranean-Diet Score
Death from coronary heart disease:
HR = 0.67 (95% CI: 0.47–0.94) for a Two-Point Increase in the Mediterranean-Diet Score
Death from cancer:
HR = 0.76 (95% CI: 0.59–0.98) for a Two-Point Increase in the Mediterranean-Diet Score
|
|
Estruch, 2013, [86]
|
Parallel-group, multicentre, randomized
trial
|
1050 men and 1493 women with MD with EVOO
1128 men and 1326 women with MD with nuts
987 men and 1463 women with Control Diet
|
Myocardial infarction, stroke, and death from cardiovascular causes:
HR = 0.70 (95% CI: 0.54–0.92, p = 0.01) for MD with EVOO vs. Control Diet
HR = 0.72 (95% CI: 0.54–0.96, p = 0.03) for MD with Nuts vs. Control Diet
Death from any cause:
HR = 0.82 (95% CI: 0.64–1.07, p = 0.15) for MD with EVOO vs. Control Diet
HR = 0.97 (95% CI: 0.74–1.26, p = 0.82) for MD with Nuts vs. Control Diet
|
|
Estruch, 2018, [87]
|
Parallel-group, multicentre, randomized
trial
|
1050 men and 1493 women with MD with EVOO
1128 men and 1326 women with MD with nuts
987 men and 1463 women with Control Diet
|
Myocardial infarction:
HR = 0.82 (95% CI: 0.52–1.30) for MD with EVOO vs. Control Diet
HR = 0.76 (95% CI: 0.47–1.25) for MD with Nuts vs. Control Diet
Stroke:
HR = 0.65 (95% CI: 0.44–0.95) for MD with EVOO vs. Control Diet
HR = 0.54 (95% CI: 0.35–0.82) for MD with Nuts vs. Control Diet
Death from cardiovascular causes:
HR = 0.62 (95% CI: 0.36–1.06) for MD with EVOO vs. Control Diet
HR = 1.02 (95% CI: 0.63–1.67) for MD with Nuts vs. Control Diet
Death from any cause:
HR = 0.90 (95% CI: 0.69–1.18) for MD with EVOO vs. Control Diet
HR = 1.12 (95% CI: 0.86–1.47) for MD with Nuts vs. Control Diet
|
|
Sofi, 2008, [88]
|
Meta-analysis of prospective cohort studies
|
1,574,299 subjects from 12 studies
|
Mortality from cardiovascular diseases:
RR = 0.91 (95% CI: 0.87–0.95)
Mortality from any cause:
RR = 0.91 (95% CI: 0.89–0.94
Mortality from cancer:
RR = 0.94 (95% CI: 0.92–0.96)
Incidence of Parkinson’s disease and Alzheimer’s disease:
RR = 0.87 (95% CI: 0.80–0.96)
|
|
Sofi, 2010, [89]
|
Meta-analysis of prospective cohort studies
|
508,393 subjects from 7 studies
|
Mortality from cardiovascular diseases:
RR = 0.90 (95% CI: 0.87–0.93)
Mortality from any cause:
RR = 0.92 (95% CI: 0.90–0.94)
Mortality from cancer:
RR = 0.94 (95% CI: 0.92–0.96)
Incidence of neurodegenerative disease:
RR = 0.87 (95% CI: 0.81–0.94)
|
|
Kromhout, 2018, [92]
|
Prospective Cohort Study
|
12,763 subjects from 16 cohorts of the Seven Countries Study
|
Mortality from cardiovascular diseases:
Inverse association between consumption of cereals, vegetables, legumes, and alcohol and long-term CHD mortality rates (r = −0.52 to −0.62)
Positive association between consumption of hard fat plus sweet products, animal foods except fish, and long-term CHD mortality rates (r = 0.68 to 0.84)
|
This entry is adapted from the peer-reviewed paper 10.3390/nu12010035