Several studies have reported higher OS in post-mortem ASD brain samples, compared to controls by means of decreased GSH levels, oxidative damage to proteins, lipids, and nucleic acids, as well as an altered activity of enzymes involved in redox balance, such as superoxide dismutase, glutathione peroxidase, glutathione-S-transferase, and glutamate cysteine ligase
[13][15]. Other studies focused their attention on erythrocytes (RBC) from ASD patients
[14][16]. Considering RBC shape, energy metabolism, and the composition of their plasma membrane-cytoskeletal network, they are thought to be sensible indicators of OS and, therefore, have been proposed as a potential biomarker in ASD. To present date, conflicting results have been reported in the literature
[15][16][17][17,18,19]. In fact, while Ciccoli et al. reported multiple shape anomalies and β-actin alterations in RBC from ASD subjects
[15][17], Giacometti et al. confirmed, through hyperspectral microscopy, neither the presence of shape alterations at a large extent
[16][18] nor β-actin content differences between RBC from ASD children and matched controls
[16][18]. In another
respape
archersr by our group
paper, they, we showed deep alterations in the erythrocyte lipid moiety, which, in turn, caused a decrease in Na
+/K
+-ATPase activity
[16][18], as also previously described
[18][20]. These modifications, occurring in blood cells, could be seen as a peripheral window on the events occurring in the central nervous system (CNS).
3. Endogenous Enzymatic Defense
In a healthy organism, in every tissue, there is a redox balance. Cells produce free radicals, but they also present an antioxidant system, made up of enzymatic and nonenzymatic antioxidants. At the end of mitochondria respiration, oxygen is the final acceptor of electrons from the electron transport chain (ETC) to produce water; however, a small fraction of electrons combines with oxygen to form the superoxide anion, which can give rise to other ROS and react with NO to form reactive nitrogen species, as mentioned before. O
2−· is produced by mitochondrial pathways and is converted to H
2O
2 by superoxide dismutase (SOD). In turn, H
2O
2 is converted to O
2 and H
2O by catalase (CAT) in peroxisomes or to H
2O via the activity of glutathione peroxidase (GPX) and oxidation of glutathione (GSH) to its disulfide form in the cytosol. So, O
2−· and H
2O
2 can create hydroxyl radicals that are able to damage DNA, proteins, and lipids when present at aberrantly high levels
[19][21].
Data present in the literature show that SOD activity in erythrocytes is higher in ASD children, in comparison with the controls
[20][22]. An increased SOD level is considered a compensatory mechanism to counteract the cell damage caused by oxidative stress within the brain. In addition, CAT activity is reduced in erythrocytes, at variance from its plasmatic activity, which is unchanged
[21][23].
In the last years, much attention has been devoted to glutathione peroxidase 1 (Gpx1), a seleno-enzyme that converts H
2O
2 into H
2O through the oxidation of GSH to oxidized glutathione (GSSG)
[22][24]. Gpx4 is also present in the mitochondria; its preferred substrates are lipid hydroperoxides, which are formed as a result of oxidative damage to the membrane phospholipids. In addition, mitochondria contain the peroxiredoxins (Prx) 3 and 5, along with their thioredoxin (Trx) 2 enzyme partner. Prx3 is exclusive to mitochondria, while Prx5 is also present in peroxisomes, in the cytosol, and in the nucleus. Prxs oxidize H
2O
2 to cysteine–SOH, which reacts with another cysteine to form H
2O and a disulfide bond. The disulfide bonds are reduced by Trx2, which cycles from a reduced to an oxidized state. Trx2 is then reduced by thioredoxin reductase (TrxR) at the expense of NADPH oxidation. These enzymes are important for maintaining the redox environment of the mitochondria and for responding to oxidative stress imposed by external environmental factors
[23][25]. Impairment of these antioxidant enzymes result in supraphysiological ROS production that can cause developmental neurotoxicity (DNT). The increased production of ROS both centrally (in the brain) and peripherally (in the plasma) may, in turn, result in the reduction of brain cell number, leading to autism pathology and apoptosis
[24][26].
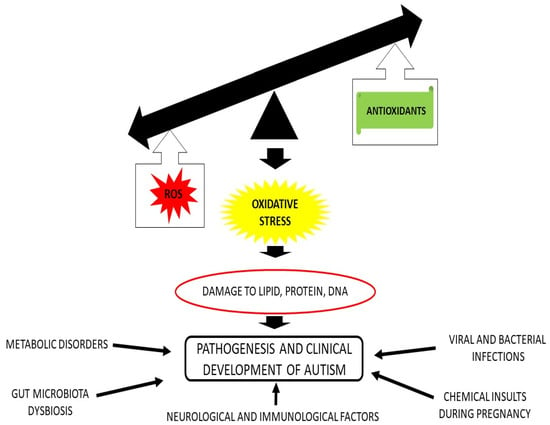
As already stated, oxidative stress is involved in the pathogenesis of ASD, since subjects suffering from ASD display an excessive ROS production, a decreased antioxidant capacity, and a mitochondrial dysfunction. All of these physiological abnormalities have the potential to cause oxidative stress, which, in turn, can cause epigenetic dysregulation, neuro-inflammation, cerebral injury, and neural cell dysfunction, which finally leads to aberrant development of CNS networks and, ultimately, to ASD
[24][26].
A recent paper by
researcheour
s group focused the attention on GPx1 enzyme. In particular, a possible correlation between the GCG repeat polymorphism present in the GPx1 gene encoding a region of five to seven alanine residues, denominated as ALA5, ALA6, and ALA7, was investigated
[25][27].
RWe
searchers showed reduced enzyme activity for GPx1 molecules carrying ALA5 sequence, followed by GPx1 expressing ALA6 and ALA7; these differences are due to the modification in the secondary and tertiary structures present in the three enzyme variants. It is of note that the ALA5 variant is not only the most frequent polymorphism among ASD subjects and unaffected family members, but also revealed the lower enzymatic activity, compared to that of the other variants tested
[25][27]. In their paper, Ming et coll. showed that, among all recruited subjects (probands and parents), there was a transmission disequilibrium of the three ALA alleles. In particular, GPX1 expressing ALA6 was significantly under-transmitted from parents to probands diagnosed with ASD, thus suggesting that owning this allele may be protecting for autistic disorder
[26][28]. Former studies, on the ALA6 allele, described a link with different human diseases, even though it was with controversial results. Specifically, it was found that ALA6 variant has an increased frequency in prostate cancer, without any association with prostate cancer risk
[27][29]. In another paper, in subjects experiencing lung tumors, the presence of ALA6 polymorphism may be protective against oxidative stress
[28][30], in accordance with
our
esearchers' study. Later on, a significant association between the GPX1, Ala6, and Ala7 alleles and squamous cell carcinoma development was proposed
[29][31]. In addition, in another paper, having one or two ALA6 alleles seemed to be related to a modest increased risk of coronary artery disease
[30][32]. In this regard, further insightful evidence and clinical reports are necessary, since GPx1 activity is only a part of a multifaceted pathway in which other proteins and enzymes are involved
[25][27].
3.1. Polymorphism in Endogenous Enzymes
As described in the previous paragraph, in addition to GPX1, SOD is a powerful enzyme devoted to the conversion of O
2−·, generated inside the cell, into H
2O
2. Few contradicting reports on the involvement of the SOD enzymes in ASD are present in the literature. Some of them describe an increased activity
[31][33], others show decreased activity
[32][34], and some show no significant change
[33][35] in the SOD enzyme activity. Kovač et coll.
[34][36] first hypothesized that the connection between the variation in SOD activity and the etiology of ASD might be due to the presence of SOD genetic variants that, in turn, could influence enzyme activity. Such an assumption was confirmed by the discovery of a significant association between the noncoding genetic variants in the SOD1 gene (rs2234694 and rs36233090) and ASD etiology.
Another important class of isoenzymes, responsible for the formation of the free radical NO, is the nitric oxide synthase (NOS) family. Three NOS isoforms, named NOS1 (neuronal NOS), NOS2 (inducible NOS), and NOS3 (endothelial NOS) have been well-defined
[35][37]. Since their discovery, many single nucleotide polymorphism (SNPs) of the NOS gene family have been described, some of which are associated with mental disorders and neurological diseases/conditions
[36][38]. Regarding ASD, in their family-based association study of ASD in a Korean cohort, Kim and coll.
[37][39] identified a significant, even though weak, association between SNPs in NOS2 gene and ASD. In their conclusions, the authors stated that many other research studies, in a larger population or other ethnic groups, are warranted.
Taken together, all these data indicate that variations in the expression of genes involved in the generation of ROS and diminution of antioxidant defense might have a significant impact on the development of the clinical symptoms of ASD. In fact, as stated elsewhere, OS may be responsible of different undesirable processes, ranging from oxidation of lipids, proteins, and DNA
[19][21], up to increase of inflammation, which, in turn, can cause brain tissue damage and alteration of the immune response
[38][40], leading to the clinical symptoms of ASD.
3.2. Endogenous Non Enzymatic Antioxidants
Besides antioxidant enzymes, eukaryotic cells possess many endogenous antioxidants acting against ROS outbreak. Among all, GSH is the main intracellular thiol used for defense against ROS, electrophiles, and xenobiotics. In fact, it can be oxidized by ROS, with the formation of the GSSG dimer, which, in turn, can be reduced by the action of glutathione reductase (GR), keeping the redox state of the cell
[23][25]. Genetic variations in glutathione pathways have been observed in ASD, and they have been correlated to ASD behaviors
[26][39][28,41]. Therefore, GSH has been indicated as a biomarker of ASD oxidative stress, since its levels in ASD patients have been reported to be variable, as some studies have reported higher levels of GSH than healthy controls, while other studies have shown lower levels
[40][42]. Nevertheless, a very recent meta-analysis highlighted that blood GSH and total glutathione (tGSH) levels are lower in ASD subjects, compared to controls
[41][43]. In another paper, Rose et al. described a negative association between the GSH/GSSG ratio and oxidative protein and DNA damage. This might suggest that diminished glutathione redox capacity in the autistic brain may have significant consequences, in terms of augmented mitochondrial superoxide production and a chronic inflammatory condition
[42][44].
Another important endogenous antioxidant is the coenzyme Q10 (CoQ10), a lipophilic molecule consisting of a benzoquinone ring with 10 isoprene units in the side chain, also known as ubiquinone. It is a very fascinating compound because, although similar in structure to some vitamins (eg, vitamin K), CoQ10 is not a vitamin, since it is synthesized in the body, whereas vitamins must be obtained from the diet. CoQ10 is an electron mobile carrier for mitochondrial enzyme complexes involved in oxidative phosphorylation in the production of adenosine triphosphate (ATP), but it also serves as a potent antioxidant or free radical scavenger
[43][45]. In its reduced form, ubiquinol, it is itself, is a potent lipophilic antioxidant and can recycle and regenerate other antioxidants in the body. Since CoQ10 is synthesized in all tissues, the body is not normally dependent on its exogenous supplies. However, endogenous biosynthesis tends to decline with age and following statin use. Moreover, tissue CoQ10 levels might be lowered in several CNS pathological scenarios, such as traumatic brain injury, and it has been suggested that, in these circumstances, CoQ10 could be supplemented to restore normal levels
[44][46]. Furthermore, an early study found a role for CoQ10 in autism. In fact, given the favourable response by ubiquinol supplementation in children with autism, the authors hypothesized that a CoQ10-dependent redox system might affect porin channel function
[45][47]. Based on these results, some authors suggested the use of CoQ10 in many brain disorders, other than ASD, comprising Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, Amyotrophic lateral sclerosis, multiple sclerosis, Friedreich’s ataxia, epilepsy, depression, and bipolar disorder, which are all associated with mitochondrial dysfunction
[43][45].
4. Exogenous Antioxidants
Exogenous antioxidants obtained from the diet are also key factors to prevent, or at least reduce, oxidative stress in humans, since particular foods contain molecules with remarkable antioxidant properties. These compounds might, therefore, protect cellular macromolecules from damage
[46][48]. There is a growing scientific consensus about a possible role of dietary agents, mainly antioxidant-containing foods, in reducing symptom severity and improving behavioural impairment in children with autism
[47][49]. Among exogenous antioxidants,
we includ
inge ascorbic acid (vitamin C), which scavenges hydroxyl and superoxide radical anion, tocopherols, and tocotrienols (vitamin E), which is involved in limiting lipid peroxidation of cell membranes and phenolic antioxidants, such as stilbene derivatives (resveratrol, phenolic acids, and flavonoids), selenium, zinc, and many more
[48][50].
Among all vitamins, vitamin C (VC) is considered one of the most studied and potent antioxidants, and it can be found in many plant cell types, organelles, and in the apoplast. Being the main hydrophilic ROS-detoxifying compound, VC is able to donate electrons in a wide range of enzymatic and non-enzymatic reactions in the aqueous phase. Thanks to its structure, VC can completely scavenge superoxide, hydroxyl radicals, and singlet oxygen and reduce H
2O
2 to water via ascorbate peroxidase reaction. In addition, VC regenerates vitamin E (VE) from its radical (tocopheroxyl, TOH·) providing membrane protection
[49][51].
On the other hand, VE, which exists in eight different isoforms (including α-, β-, γ-, δ- tocopherol, and the corresponding tocotrienol), is synthesized only by plants and algae and represents the most important lipid phase antioxidant. All isoforms differ in their substituents, and α-tocopherol, with its three methyl substituents, possesses the highest antioxidant activity among all tocopherols
[50][52]. VE is classified as a chain-breaking antioxidant, since it can repair oxidizing radicals directly, thus preventing lipid peroxidation due to PUFA oxidation. This typical reaction occurs in the membrane–water interphase, where VE is transformed into TOH·. Regeneration of the reduced form can be achieved by VC, GSH, or CoQ10. In addition to these mechanisms, tocopherols can act either as scavengers of oxygen radicals or as physical deactivators of singlet oxygen
[49][51]. Differently from lipid-soluble VE, VC is not stored within the body, and since it is not endogenously formed in humans, it is promptly excreted in the urine, thus requiring a continuous supplementation from the diet. Hence, VC plasma concentration is related to its dietary intake, having a half-life of 12–24 h
[51][53]. Different recent studies have shown lower VC levels in the blood of ASD children, compared to healthy controls
[52][53][54,55]. In accordance with these studies, it has also been described that blood VE levels are reduced in autistic children, and they are associated with ASD-like behaviors in the same subject
[54][56].
Although lacking antioxidant properties, various vitamins of the B group, such as vitamin B6 (pyridoxine), vitamin B9 (folic acid), and vitamin B12 (cobalamin), play important roles in the development, differentiation, and functioning of the CNS, as they are implicated in the methionine-homocysteine pathway. Metabolically active forms of vitamin B12, as well as folate, serve as essential cofactors for two key enzymes important for DNA synthesis, serine synthesis, for the regeneration of methionine for protein synthesis and methylation, and for the prevention of homocysteine accumulation. Since vitamin B12 is only synthesized by specific bacteria, humans can obtain it from animal source foods, such as dairy products, meat, eggs, fish, and shellfish
[55][57]. Studies from the literature have pointed out that vitamin B12 levels are decreased in ASD children, compared to healthy children
[56][58]. Such deficiency causes a decrease in homocysteine re-methylation, hence resulting in increased homocysteine levels, as well as in DNA hypomethylation, affecting the development of the CNS
[57][59]. Vitamin B deficiency can be due to a lack of nutrients, poor absorption, or intestinal disorders, where the gut microbiota plays a crucial role, since it synthesizes essential dietary vitamins and cofactors
[24][26]. In addition, a concomitant deficiency of vitamins, B6, B9, and B12 was found in a great number of autistic subjects that, in turn, may cause DNA methylation deficiency. This deficiency might be explained as related to intestinal dysbiosis, which is the main cause of vitamin intestinal malabsorption
[58][60]. In order to counteract this situation, different clinical trials have been conducted, with the aim of restoring vitamin E/vitamin C/glutathione antioxidant network in ASD individuals
[59][61]. They observed a positive correlation between changes in behavior and GSH/GSSG ratio. Thus, according to scientific evidences, ASD-like symptoms may be improved by targeting the vitamin E/vitamin C/glutathione rheostat. Although the clinical effectiveness of nutritional interventions heightening vitamin E and vitamin C levels, both present within foods, in restoring an optimal functionality of the antioxidant network has not yet been investigated on a large scale, current evidence supports a putative role
[47][49].
Among exogenous antioxidants, found in a wide variety of cultivated plants and derived foods, polyphenols are a group of water-soluble secondary metabolites (flavonoids, tannins, hydroxycinnamate esters, and lignin). Their antioxidant properties arise from their high reactivity as hydrogen or electron donors and from their ability to stabilize and delocalize the unpaired electron, as well as to chelate transition metal ions
[60][62]. Another mechanism related to the antioxidant properties of polyphenols is the ability of flavonoids to alter peroxidation kinetics by modification of the lipid packing order and to modulate membrane fluidity
[61][63]. These changes could sterically hinder the diffusion of free radicals and restrict oxidative reactions.
To date, very few studies have explored the possible benefits of dietary polyphenols in ASD patients. Some of these were performed in order to alleviate ASD symptoms, and they have demonstrated that dietary polyphenols, including resveratrol, could be promising molecules, since they are able to modulate mitochondrial activity and dysfunction, usually present in individuals with ASD
[62][64]. In addition, resveratrol was recognized to suppress ROS production inside mitochondria in stimulated primary hippocampal astrocytes
[63][64][65,66]. It is important to take into account that not all polyphenols can cross the blood–brain barrier, and for this reason, they can display a low bioavailability in the brain, despite being highly absorbed from the intestinal lumen. Therefore, they may putatively alter microbiota composition, which is of great importance for ASD, usually characterized by alterations in the microbiota–gut–brain axis
[65][67]. On the whole, current evidence suggests that polyphenols may represent a class of natural compounds, with a potential utility in treating ASD symptoms
[66][68].