In vitro culture of endothelial cells to form capillary-like networks is essential in tissue engineering. Vascular endothelial growth factor (VEGF) is one of the primary signal proteins stimulating blood vessel formation. This growth factor can be soluble in the medium or protein-bound to the substrate. However, less attention has been paid to distinguishing the specific stimulations by soluble and bound VEGF. We conducted a series of experiments to explore the respective effects of these two VEGF forms. An in-house synthesized biogel comprising a definite concentration of collagen and fibronectin was designed to cultivate human umbilical vein endothelial cells to form the capillary-like network. Collagen served as the primary substrate for cell attachment. Fibronectin provided the surface to bind soluble VEGF in the culture medium to create the bound VEGF. The experiment of adding VEGF-blocking-peptide was conducted to prevent the formation of VEGF bound to the fibronectin domains, to distinguish the respective effects of the soluble and bound VEGF. With the in-house biogel of definite components, we were able to clarify the different roles of soluble and bound VEGF. The results indicated that the soluble VEGF promptly induced the cells to change from round to elongated shape, which contributed to forming network cords. Simultaneously, the bound VEGF provided long-term stimulation, causing the cells to migrate and differentiate into the final capillary-like network.
1. Introduction
The effect of VEGF on vasculogenesis is the key to cultivate a capillary network in the engineered tissue. In vivo, VEGF could be soluble in the medium or protein-bound
[[1],[2],[3]]; however, how the soluble and bound forms of VEGF regulate the network forming is still not completely clear. The biogel-bound VEGF (abbreviated as the bound VEGF afterward) dominates angiogenesis and vasculogenesis by providing the prolonged VEGFR2 activation and subsequent enhancement of the phosphorylated pathway
[[4],[5],[6]]. Additionally, the synergy of integrin and VEGFR2 on endothelial cell membranes has been discovered
[[7]]. With the integrin-binding and growth factor-binding domains located closely enough on fibronectin, the chemical stimulation for endothelial cell migration is believed to be further enhanced. However, to date, a correct understanding of soluble or bound VEGF binding phenomena and their corresponding function is still unclear.
2. Analysis on Results
2.1. HUVECs Cultivation on Type I Collagen and Fibronectin
The primary purpose of this study is to explore the respective effect of soluble and bound VEGF on endothelial cells. Therefore, instead of using the commercial matrix (such as Matrigel
®), which already contains growth factors, pure collagen type I and fibronectin were used to make the in-house biogel with a definite combination. Type I collagen as the primary substrate provided the endothelial cells with attachment sites when the cells moved. Fibronectin attached to the collagen surface provided cell attachment and binding sites for soluble VEGF initially contained in the culture medium to form bound VEGF. The cell suspension was added to a 12-well plate coated with collagen with or without fibronectin. The initial number of endothelial cells was 80,000 cells per well, and the cell distribution images were recorded every 6 h.
The endothelial cells were cultivated under four conditions, considering whether fibronectin was coated on the collagen and whether VEGF was added to the culture medium. The endothelial cells in the four cases formed the network to different extents. The results are as shown in
Figure 12. In the two cases with no VEGF but with or without fibronectin addition (−F−V and +F−V), HUVECs started the morphological change from round to elongated shape at about 6 h, and the change became evident at 12 h. The cells in these two cases were found to undergo a rapid cell death process that the number of adherent endothelial cells continually decreased. At 24 h of incubation, only a small number of cells remained in the sight field. It is worth mentioning that the cells could express the morphological change even without VEGF added.
Figure 12. HUVECs morphological forms and cellular networks in different cell culture conditions: without fibronectin (−F), with fibronectin (+F), without VEGF (−V), and with VEGF (+V). Black arrows in the enlarged photos indicate the cells with morphological change into the elongated shape, while white arrows indicate the cells still in the original cobblestone shape.
Under the culture condition without fibronectin but with VEGF (−F+V), many cells with morphological changes appeared at 6 h. However, the cell network could not ultimately form afterward. In the +F+V case, the beginning of morphological change occurred a bit later than in the –F+V case. The originally round-shaped cells became elongated, and the dividing boundary for adjacent cells became blurred at 12 h. These characteristics were more evident subsequently, which shows that VEGF stimulated the endothelial cells continuously. The cell-free area in the visual field became apparent as a result of cell aggregation. The cell colony evolved in the direction of forming the capillary-like network.
2.2. Effects of Soluble and Bound VEGF
Comparison between the –F+V and +F+V cases, as shown in
Figure 12, reveals that the endothelial cells begin morphological changes earlier; however, a cell network could not form afterward if the biogel had no fibronectin. In contrast, although the cell commenced morphological change somewhat later in +F+V, the cells continued to migrate and aggregate into a cellular network. This result indicated that soluble and bound VEGF might have different effects on cell behavior. However, in addition to binding VEGF, fibronectin could bind cells through its integrin-binding domain. To clarify whether it was the cell-binding or VEGF-binding to the fibronectin responsible for the different cell behaviors, we conducted a series of experiments using the synthetic VEGF-blocking-peptide (VBP). The sequences of VBP were designed according to the fibronectin-binding domain of VEGF (exon-7). This peptide was added to the fibronectin-coating wells before adding the HUVEC suspension to shield the growth factor-binding domain of fibronectin. Because the bound VEGF was prevented from forming, the added VEGF would remain in the soluble form.
The VPB shielding effect was assessed using fluorescence immunoassay, where a FITC-conjugated anti-VEGF antibody was used to monitor the VEGF. Results are as shown in
Figure 23 for four different cell culture conditions: without VBP (-P), with VBP (+P), without VEGF (-V), and with VEGF (+V). The darker fluorescence response in the case of +P+V than in –P+V indicated that VBP could avoid VEGF from biding fibronectin. The two subsets of -P-V and +P-V showed that even in the cases with no adding VEGF in the culture medium, the fluorescence response could still be detected, albeit the responses were weaker than the counterpart cases of -P+V and +P+V, respectively. The results indicated that HUVECS could secrete VEGF. This autocrine signaling explained why HUVECs presented a morphological change in the cases without adding VEGF from outside. However, because the cells experienced rapid cell death in the two cases without VEGF added, adding VEGF was essential to cell viability to prevent the cell un-attachment and cell death during the network developing process.
Figure 23. Bound VEGF location after 6 h of incubation in different cell culture conditions: without VBP (−P), with VBP (+P), without VEGF (−V), and with VEGF (+V). A fluorescent signal from the primary antibody against VEGF was detected on the cell membrane and biogel surface. The VBP shielding effect avoided VEGF from binding biogel resulting in the darker VEGF field in both the +P−V and +P+V cases.
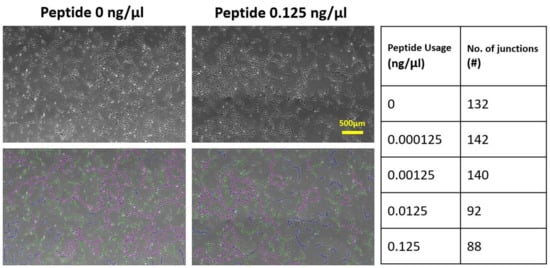
We further tested the VBP effect on cells by changing dose usage. The number of junctions quantitatively indicated the integrity and complexity of the capillary-like network. The results are displayed in
Figure 34, which shows HUVECs could differentiate into a network and established an average of 140 junctions per view field for the cases with no or adding a small amount of VBP. The junction number dramatically decreased with the peptide concentration larger than 0.0125 ng/μL, but the cells kept attaching to the biogel. The results showed that the usage of VBP interfered with the regulating effect of VEGF and affected the integrity of network structure but not the cell adhesion.
Figure 34. The effect of VBP dosage on the cell network integrity. The average number of network junctions per view field served as a quantitative indicator of network integrity. Cell cords recognized by ImageJ software are shown, where the pink lines present the segments (connections between two junctions), the green lines the branches (connections between junction and node), and the blue lines the isolated elements (connections between two nodes). Data were collected after 24 h of cell culture and averaged with three repeated experiments.
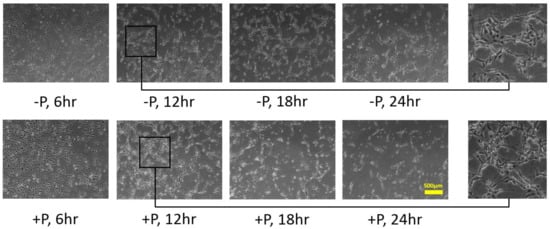
Based on the optimum culturing environment (+F+V), as shown in
Figure 12, we tested on HUVECs the stimulating effect of soluble and bound VEGF, respectively, by adding 400 μL DPBS per well with 0.125 ng/μL dissolved VBP. The cell culture images are shown in
Figure 45. In the -P situation, i.e., without VBP addition, HUVECs showed the change in morphology and subsequently formed the capillary-like network. In the +P situation, i.e., with VBP addition, HUVECs could not ultimately create the cell network, albeit the cells presenting morphological change earlier than the -P case.
Figure 45. HUVECs distribution with and without VBP addition. In the -P case, which means without the VBP addition, HUVECs showed the change in morphology at 12 h with some cells remaining in the cobblestone shape and subsequently formed the capillary-like networks at 18 h. In the +P case, which means with the VBP addition, most HUVECs showed the morphological change at 12 h; however, the network could not form afterward.
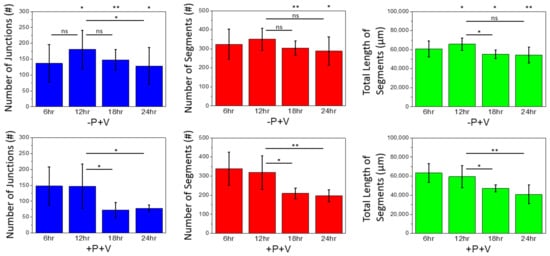
More details about the network integrity and complexity are summarized in
Figure 56. The number of junctions, the number of segments, and the total segment length were measured as representative indicators. If no VBP was added (−P+V), these three values remain high for all the observation times, indicating that a cell network was developed to a high degree. In contrast, with VBP added (+P+V), these three values decreased significantly after 18 h, indicating no cell network could ultimately form with VBP added. Because adding VBP (+P) prevented VEGF binding onto fibronectin, the added VEGF would remain soluble. The soluble VEGF was shown incapable of sustaining the network formation.
Figure 56. Junction number, segment number, and total segment length of the capillary-like network. The higher value of these three indicators, the more integrity of the network. All experiments were repeated five times. Data are presented as mean SD. The Student’s t-test was performed for statistical evaluation: * = p < 0.05, ** = p < 0.01, and ns = non-significant. The asterisk above the -P+V charts presents the p-value between the −P+V and +P+V cases at the same observation time.
3. Current Insights
Previous experiments primarily focused on the VEGF amount added to the culture environment, lacking the discussion on the two states of VEGF
[[8],[9]]. Less attention was paid to telling the different effects of soluble and bound VEGF. The hypothesis that the formation of capillary-like networks is only regulated by the bound VEGF cannot fit all experimental data
[[10]]. In theoretical modeling, on the other hand, capillary-like networks can be formed by using both soluble-VEGF-based or bound-VEGF-based mathematical models, and a consensus is still absent
[[11],[12],[13]]. Moreover, although many in vitro capillaries culturing experiments have been successful, problems remained in the in vivo trials. Once the artificial scaffold and tissue are transplanted into the human body, the VEGF binding to the scaffold will gradually be consumed or decomposed over time. How to regularly provide an appropriate amount of VEGF becomes a difficult challenge
[[14],[15]]. A high initial dosage strategy is not adopted because the excessively high VEGF concentration will cause excessive proliferation of capillaries and potential carcinogenic risks
[[16]]. Transplanting biomaterials, such as fibronectin, that can initially capture and gradually release VEGF, seems feasible, while it is essential to precisely predict the dynamic balance between soluble and bound VEGF
[[17],[18]].
Previous experiments showed that parts of the cells with morphological change to elongated shape are connected end-to-end to form the cords in the capillary-like network, while those without morphological change aggregate and create a cobblestone arrangement
[[9],[19]]. The same phenomena were found in our results (+F+V in
Figure 12 and -P in
Figure 45). Because the animal-extracted materials such as Matrigel and ECGS used in literature are originally rich in VEGF, previous studies lacked the discussion on the respective effects of soluble and bound form. We filled this gap by building a VEGF-free environment, which allowed us to explore the VEGF effects. We chose the type I collagen as substrate ingredient, which had been proven beneficial for HUVECs survival and biogel remodeling
[[8],[20]], and treated with the thin fibronectin layer coated on collagen surface, which provided VEGF-binding areas, and further enhanced HUVECs attachment and morphology
[[7],[17]]. By comparing the cases with and without adding VEGF, we discussed the different roles soluble and bound VEGF played in forming capillary-like networks.
The exogenous VEGF tests on HUVECs showed that more cells underwent a morphological change in the −F+V condition within 12 h of incubation than the standard +F+V case. Since in the −F+V case there was virtually no VEGF binding to fibronectin, the added VEGF remains in the soluble form; the results indicated a higher concentration of soluble VEGF induced the cell morphological change earlier. The primary response of endothelial cells to the stimulation of soluble VEGF was the shape change from round to elongated shape.
The experiment of adding VEGF-blocking-peptide (VBP) to prevent the formation of bound VEGF further confirmed the respective effects of soluble and bound VEGFs. Adding VBP resulted in a higher concentration of soluble VEGF in the culture medium than not adding VBP. Therefore, we could confirm that soluble VEGF mainly stimulated the endothelial cells to change from a round to elongated form, which might benefit the center-hollowed tube structure development
[[21]]. On the other hand, the bound VEGF mainly stimulated the endothelial cells to migrate and ultimately formed a high integrity network.
In summary, the soluble and bound VEGF had different vital functions in inducing endothelial cells to differentiate into a capillary-like network. The soluble VEGF dissolved in the culture medium activated the cell morphogenesis quickly. The cells became elongated, which was conducive to the formation of network cords. Simultaneously, the VEGF binding to the biogel provided a slow and continuous chemical stimulation, guiding the cells to move and aggregate, forming a capillary-like structure with complete integrity.