2. Preparation of Phenolic Solutions
HT and OL were dissolved in culture medium (DMEM/F-12 + GlutaMax, without any supplement), and freeze dried OP extracts CONV, OPT1, OPT2, and OPT3 were dissolved in 0.4% EtOH in culture medium, on the same day of the experiment.
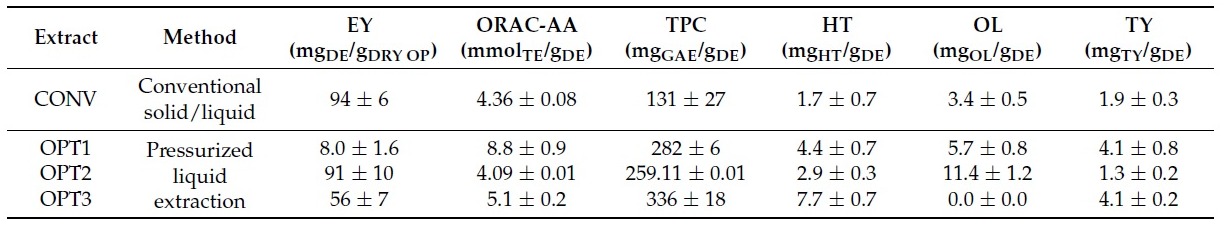
The OP extracts were prepared as previously described [38]. The CONV extract was prepared by conventional solid-liquid extraction from freeze dried OP while OPT1, OPT2 and OPT 3 were produced by pressurized liquid extraction from freeze-dried OP previously deffated by supercritical carbon dioxide extraction. Their composition is described in Table 1. The extraction conditions for OPT1, OPT2 and OPT3 were optimized by a desing of experiments [38] so that they correspond to the extract with the highest chemical antioxidant activity by ORAC assay (Oxigen Radical Absorbance Capacity assay), the highest concentration of OL, and the highest concentration of HT, TY and TPC (total phenolic concentration), respectively.
Table 1. Characterization of the conventional (CONV) and the three optimized (OPT1, OPT2, and OPT3) olive pomace (OP) extracts in terms of extraction yield (EY) (as mg of dry extract (DE)/g of DRY OP), oxygen radical absorbance capacity antioxidant activity (ORAC-AA) (as mmol of Trolox equivalents (TE)/g of DE), and total phenolic content (TPC) (as mg of gallic acid equivalents (GAE)/g of DE), as well as extract richness in Hydroxytyrosol (HT), Oleuropein (OL) and Tyrosol(TY) (as mg of compound/g of DE). The results are presented as average ± standard deviation.
3. Anti-Inflammatory Activity of Phenolic Solutions
The anti-inflammatory activity of the non-toxic concentrations of the phenolic solutions was tested on the TNF-α-induced cytokine/chemokine secretion by HCE and IM-ConjEpi cells. IL-6, IL-8, IL-1β, and IP-10 secretion was significantly stimulated by 25 ng/mL TNF-α on both cell lines. In the case of IL-1β, the stimulation was not sufficient in two experiments, while IL-17A production was not increased by the stimulus selected neither on HCE, nor on IM-ConjEpi cells (data not shown).
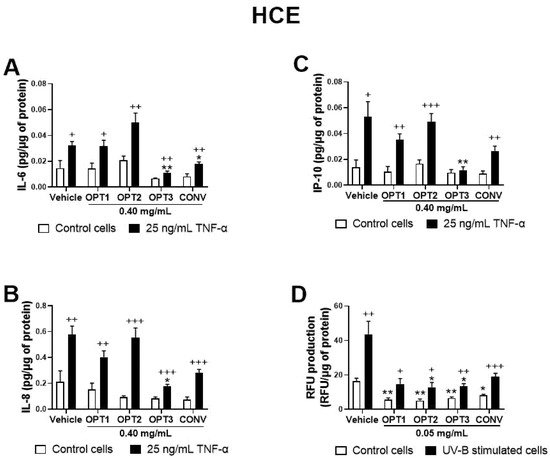
As a first preliminary screening, four OP extracts, CONV, OPT1, OPT2, and OPT3, were tested on HCE at the same concentration (0.40 mg/mL) to compare their activity. Figure 2A–C shows their effect on IL-6, IL-8, and IP-10 secretion for this cell line. IL-1β was not detected in this experiment (data not shown). CONV was able to significantly decrease IL-6 secretion by HCE cells, while optimized extract OPT3 significantly inhibited all the measured cytokines/chemokines. Moreover, the effect of OPT3 on IP-10 secretion was so strong that, in its presence, no significant differences were observed between TNF-α-stimulated and non-stimulated cells. Comparing the results among the four extracts, OPT3 demonstrated significantly higher anti-inflammatory activity compared with the rest of the extracts for IL-6 (p-value = 0.032 with OPT1; p-value = 0.014 with OPT2; p-value = 0.045 with CONV) and IL-8 (p-value = 0.026 with OPT1; p-value = 0.022 with OPT2; p-value = 0.032 with CONV). Furthermore, compared with OPT2, OPT3 had stronger inhibitory activity for IP-10 (p-value = 0.004), and CONV for IL-6 (p-value = 0.031). Therefore, according to the preliminary assay results, CONV and OPT3 were the only two extracts selected for dose-dependent studies, along with HT and OL, in both cell lines (shown in Figure 3, Figure 4, Figure 5 and Figure 6).
Figure 2. Screening of the anti-inflammatory and antioxidant activity of the conventional (CONV) and the three optimized (OPT1, OPT2, and OPT3) Olive Pomace (OP) extracts on TNF-α-induced cytokine release and UV-B-induced reactive oxygen species (ROS) production by HCE cells, respectively. For cell cytokine stimulation, cells were pre-treated with vehicle (0.4% ethanol) or 0.40 mg/mL of CONV, OPT1, OPT2, and OPT3 for 2 h. Following this, they were stimulated with 25 ng/mL TNF-α in the presence of the treatments for 24 h (A–C, black bars). Vehicle-treated-TNF-α stimulated cells and cells not stimulated with TNF-α (A–C, white bars) were used as control. IL-6, IL-8, and IP-10 were measured in cell supernatants by a multiplex bead-based array. OPT3 inhibited all measured cytokines/chemokines (A–C), while CONV significantly decreased IL-6 (A). For the UV-B-induced ROS production, cells were pre-treated with vehicle (0.4% ethanol) or 0.05 mg/mL of CONV, OPT1, OPT2 and OPT3 for 1 h. Subsequently, cells were incubated with 10 μM H2DCF-DA solution for 30 min, and then treated with the treatments and exposed to 107.25 mJ/cm2 UV-B light (D, black bars). After 1 h of incubation, intracellular fluorescence intensity was measured. Vehicle-treated-UV-B stimulated cells and cells not stimulated with UV-B (D, white bars) were used as control. OPT2 and OPT3 decreased ROS levels significantly. Data are presented as picograms (pg) of cytokine/chemokine (for cell cytokine stimulation) or relative fluorescence units (RFUs) (for ROS production) normalized to micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, ** p < 0.01, compared with vehicle-treated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells. Vehicle: 0.4% EtOH.
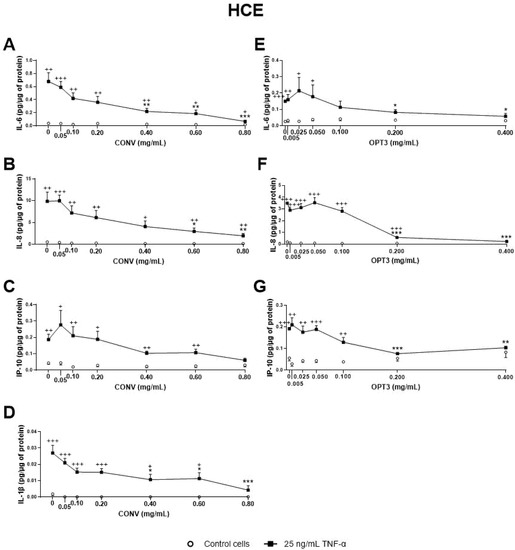
Figure 3. Effect of the conventional (CONV) and the selected optimized (OPT3) olive pomace (OP) extracts on TNF-α-induced cytokine release by HCE cells. Cells were pre-treated with CONV (0.05, 0.10, 0.20, 0.40, 0.60, and 0.80 mg/mL), OPT3 (0.005, 0.025, 0.050, 0.100, 0.200, and 0.400 mg/mL), or vehicle (0.4% ethanol) for 2 h. Following this, they were stimulated with 25 ng/mL TNF-α in the presence of the treatments for 24 h (black squares). Vehicle-treated-TNF-α stimulated cells and cells not stimulated with TNF-α (white circles) were used as control. IL-6, IL-8, IP-10, and IL-1β were measured in cell supernatants by a multiplex bead-based array. TNF-α failed to stimulate IL-1β in the experiment performed for OPT3. CONV significantly decreased IL-6 levels from 0.40 mg/mL (A), IL-8 levels from 0.60 mg/mL (B), and IL-1β levels from 0.40 mg/mL (D). For TNF- α stimulated cells, IP-10 production was not decreased significantly by CONV. Nevertheless, no significant differences were observed between stimulated and non-stimulated cells at 0.80 mg/mL (C). OPT3 inhibited IL-6 (E), IL-8 (F), and IP-10 (G) secretion from 0.200 mg/mL significantly. Data are presented as picograms (pg) of cytokine/chemokine per micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, ** p < 0.01, *** p < 0.001, compared with vehicle-treated-TNF-α stimulated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells.
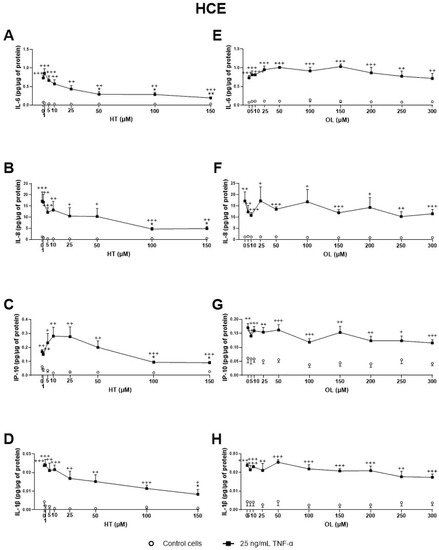
Figure 4. Effect of Hydroxytyrosol (HT) and Oleuropein (OL) on TNF-α-induced cytokine release by HCE cells. Cells were pre-treated with HT (1, 5, 10, 25, 50, and 100 μM for both cell lines, and 150 μM for HCE), OL (5, 10, 25, 50, 100, 150, 200, 250, and 300 μM) or vehicle (cell culture medium) for 2 h. Following this, they were stimulated with 25 ng/mL TNF-α in the presence of the treatments for 24 h (black squares). Vehicle-treated-TNF-a stimulated cells and cells not stimulated with TNF-α (white circles) were used as control. IL-6, IL-8, IP-10, and IL-1β were measured in cell supernatants by a multiplex bead-based array. HT significantly decreased IL-6 (A), IL-8 (Β), IP-10 (C), and IL-1β (D) secretion at 50, 100, 100, and 150 μM, respectively. OL failed to inhibit any of the cytokines/chemokines measured (E–H). Data are presented as picograms (pg) of cytokine/chemokine per micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, ** p < 0.01, compared with vehicle-treated-TNF-α stimulated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells.
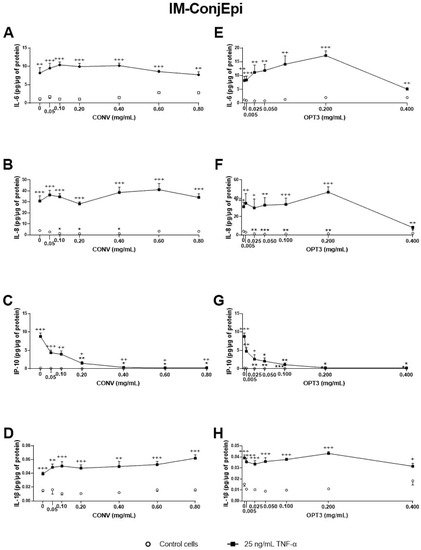
Figure 5. Effect of the conventional (CONV) and the selected optimized (OPT3) olive pomace (OP) extracts on TNF-α-induced cytokine release by IM-ConjEpi cells. Cells were pre-treated with CONV (0.05, 0.10, 0.20, 0.40, 0.60, and 0.80 mg/mL), OPT3 (0.005, 0.025, 0.050, 0.100, 0.200, and 0.400 mg/mL) or vehicle (0.4% ethanol) for 2 h. Following this, they were stimulated with 25 ng/mL TNF-α in the presence of the treatments for 24 h (black squares). Vehicle-treated-TNF-α stimulated cells and cells not stimulated with TNF-α (white circles) were used as control. IL-6, IL-8, IP-10, and IL-1β were measured in cell supernatants by a multiplex bead-based array. CONV and OPT3 reduced IP-10 levels at 0.20 mg/mL (C) and 0.05 mg/mL (G), respectively. IL-8 levels at baseline cells were decreased by CONV and OPT3 at 0.10 mg/mL (B) and 0.025 mg/mL (F), respectively. No significant inhibition was observed for IL-6 and IL-1β, neither by CONV (A,D, respectively) nor by OPT3 (E,H, respectively). Data are presented as picograms (pg) of cytokine/chemokine per micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, ** p < 0.01, *** p < 0.001, compared with vehicle-treated-TNF-α stimulated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells.
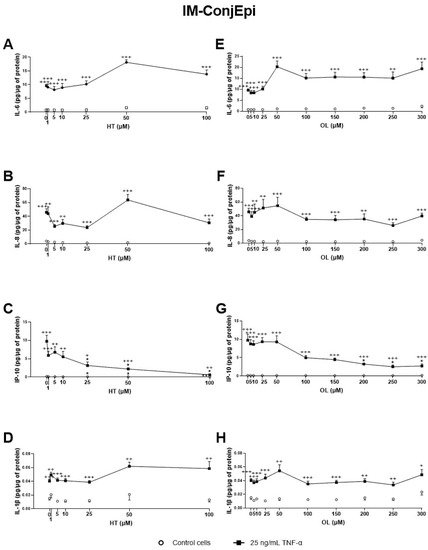
Figure 6. Effect of Hydroxytyrosol (HT) and Oleuropein (OL) on TNF-α-induced cytokine release by IM-ConjEpi cells. Cells were pre-treated with HT (1, 5, 10, 25, 50, and 100 μM), OL (5, 10, 25, 50, 100, 150, 200, 250, and 300 μM), or vehicle (cell culture medium) for 2 h. Following this, they were stimulated with 25 ng/mL TNF-α in the presence of the treatments for 24 h (black squares). Vehicle-treated-TNF-α stimulated cells and cells not stimulated with TNF-α (white circles) were used as control. IL-6, IL-8, IP-10, and IL-1β were measured in cell supernatants by a multiplex bead-based array. HT and OL reduced IP-10 levels at 25 μM (C) and 200 μM (G), respectively. No significant inhibition was observed for IL-6, IL-8, and IL-1β, neither by HT (A,B,D, respectively) nor by OL (E,F,H, respectively). Data are presented as picograms (pg) of cytokine/chemokine per micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, compared with vehicle-treated-TNF-α stimulated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells.
Regarding the OP extracts’ effect on HCE cells, as shown in Figure 3, CONV significantly decreased IL-6, IL-8, and IL-1β TNF-α stimulated secretion by cells from 0.40, 0.60, and 0.40 mg/mL, respectively (Figure 3A–D). It is also important to note that, at a concentration of 0.8 mg/mL, no statistically significant differences in IL-1β and IP-10 levels were observed between TNFα-stimulated and non-stimulated cells, demonstrating a preventive anti-inflammatory effect. On the other hand, OPT3 significantly decreased stimulated IL-6, IL-8, and IP-10 secretion by HCE cells from 0.200 mg/mL, whereas IL-1β was not detected in this experiment (Figure 3E–G). Further, OPT3 prevented IL-6, IL-8, and IP-10 production at 0.100, 0.400, and 0.200 mg/mL, respectively. Regarding pure phenolic compounds, HT significantly inhibited the secretion of IL-6, IL-8, and IP-10 from 50, 100, and 100 μM (Figure 4A–C), respectively, while only 150 μM of HT was able to decrease IL-1β levels (Figure 4D). However, OL did not demonstrate any significant anti-inflammatory effect on HCE cells (Figure 4E–H).
Additionally, the effect of CONV and OPT3 extracts and of HT and OL on cytokine secretion by IM-ConjEpi cells is shown in Figure 5; Figure 6, respectively. CONV, OPT3, HT, and OL significantly decreased IP-10 levels starting from 0.20 mg/mL (Figure 5C), 0.05 mg/mL (Figure 5G), 25 μM (Figure 6C), and 200 μM (Figure 6G), respectively. In the case of OPT3, from 0.050 mg/mL, no significant differences were observed for IP-10 levels between TNF-α- stimulated and non-stimulated cells, while from 0.025 mg/mL, IP-10 levels also decreased in non-stimulated cells (Figure 5G). The preventing anti-inflammatory effect of OPT3 was also observed for IL-8, whose levels in the baseline cells decreased significantly from 0.025 mg/mL (Figure 5F). Cytokine secretion inhibition effect in non-stimulated cells was also observed by 0.10–0.40 mg/mL of CONV for IL-8 (Figure 5B), and by HT from 25 μM for IP-10 (Figure 5C).
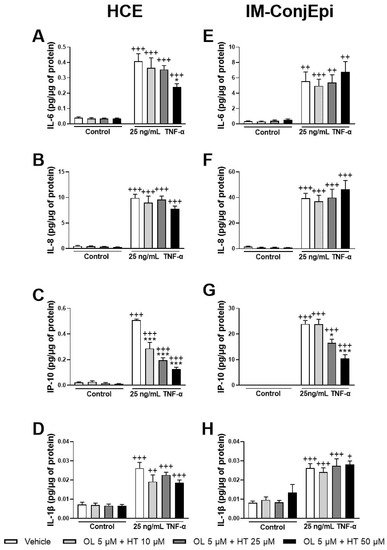
The anti-inflammatory effect of different mixtures of low concentrations of the pure phenolic compounds was also studied. The mixture of 5 μM of OL + 10 μM of HT significantly decreased TNF-α-induced IP-10 secretion by HCE cells (Figure 7C), demonstrating a strong synergistic effect. Further, 5 μM of OL combined with 50 μM of HT also inhibited IL-6 production by HCE cells (Figure 7A), and 5 μM of OL with 25 μM of HT decreased IP-10 production by IM-ConjEpi cells (Figure 7G). However, these effects were already demonstrated by HT alone (Figure 4A and Figure 6C).
Figure 7. Effect of the mixture of Hydroxytyrosol (HT) and Oleuropein (OL) on TNF-α-induced cytokine release by HCE (A–D) and IM-ConjEpi (E–H) cells. Cells were pre-treated with 5 μM of OL + 10 μM of HT, 5 μM of OL + 25 μM of HT, 5 μM of OL + 50 μM of HT, or vehicle (cell culture medium) for 2 h. Following this, they were stimulated with 25 ng/mL TNF-α in the presence of the treatments for 24 h. Vehicle-treated-TNF-α stimulated cells and cells not stimulated with TNF-α were used as control. IL-6, IL-8, IP-10, and IL-1β were measured in cell supernatants by a multiplex bead-based array. For HCE cells, 5 μM of OL + 10 μM of HT had a synergistic effect, decreasing IP-10 levels (C), whereas the decrease of IL-6 levels by 5 μM of OL + 50 μM of HT (A) can also be achieved by 50 μM HT alone. None of the mixtures tested were able to decrease IL-8 (B) or IL-1β (D) secretion significantly. For IM-ConjEpi cells, 5 μM of OL + 25 μM of HT reduce IP-10 production (G); however, this is also demonstrated by HT alone. None of the mixtures were able to inhibit IL-6 (E), IL-8 (F), or IL-1β (H) secretion significantly. Data are presented as picograms (pg) of cytokine/chemokine per micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, *** p < 0.001, compared with vehicle-treated-TNF-α stimulated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells. Vehicle: Cell culture medium.
4. Antioxidant Activity of Phenolic Solutions
The antioxidant activity of the phenolic solutions was assayed on the UV-B-induced intracellular ROS production on both ocular surface cells, selecting only the non-toxic concentrations as determined by the XTT assay. Same as for the anti-inflammatory activity, a first screening for the effect of the four different OP extracts (at the same concentration, 0.05 mg/mL) was performed on HCE cells. As shown in Figure 2D, all extracts were able to reduce ROS levels by the baseline cells, acting preventively. However, only OPT2 and OPT3 decreased ROS levels significantly in the UV-B-induced cells at the selected concentration, demonstrating a curing antioxidant effect.
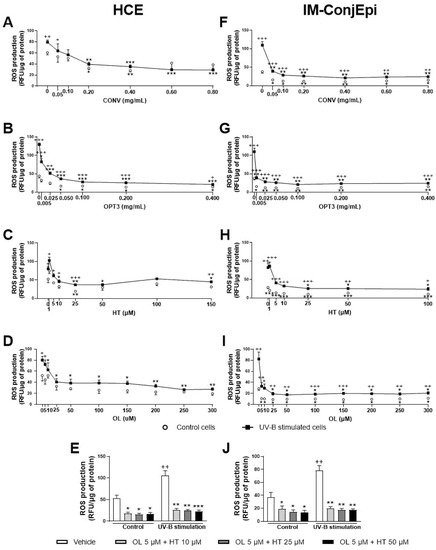
Further, dose-dependent studies were performed for the antioxidant effect of CONV and OPT3 extracts, together with HT and OL on both HCE and IM-ConjEpi cells. Figure 8 presents the antioxidant activity based on the dose-dependent studies performed for CONV, OPT3, HT, OL, and OL + HT mixtures. In the case of HCE cells, CONV and OPT3 significantly inhibited ROS production, starting from 0.20 mg/mL (Figure 8A) and 0.005 mg/mL (Figure 8B), respectively. It is important to highlight that, for CONV, no significant differences were observed between UV-B exposed and non-exposed cells from 0.10 mg/mL, while OPT3 decreased the ROS levels significantly in non-exposed cells from 0.050 mg/mL. Therefore, both extracts also demonstrate a preventive antioxidant effect. HT was able to inhibit ROS production from 10 μM (except 100 μM), while the concentrations of 50 and 100 μM prevented it (Figure 8C). OL also decreased ROS levels from 10 μM with no significant differences compared with baseline cells, demonstrating a strong curing and preventive antioxidant activity (Figure 8D).
Figure 8. Effect of Hydroxytyrosol (HT), Oleuropein (OL), and their combination, together with the conventional (CONV) and the selected optimized (OPT3) olive pomace (OP) extracts on UV-B-induced reactive oxygen species (ROS) production by HCE HCE (A–E) and IM-ConjEpi (F–J) cells. Cells were pre-treated with OL (5, 10, 25, 50, 100, 150, 200, 250, and 300 μM), HT (1, 5, 10, 25, 50, and 100 μM for both cell lines, and 150 μM for HCE), OL + HT (5 μM OL + 10 μM HT, 5 μM OL + 25 μM HT, 5 μM OL + 50 μM HT), CONV (0.05, 0.10, 0.20, 0.40, 0.60, and 0.80 mg/mL), OPT3 (0.005, 0.025, 0.050, 0.100, 0.200, and 0.400 mg/mL), or vehicle (cell culture medium for HT and OL or 0.4% ethanol for the extracts) for 1 h. Subsequently, cells were incubated with 10 μM H2DCF-DA solution for 30 min, and then treated with the treatments and exposed to 107.25 mJ/cm2 UV-B light (A–D and F–I, black squares; E,J, black bars). After 1 h of incubation, intracellular fluorescence intensity was measured. Vehicle-treated-UV-B stimulated cells and cells not stimulated with UV-B (A–D and F–I, white circles; E,J, white bars) were used as control. CONV, OPT3, HT, and OL inhibited ROS production by HCE cells at 0.20 mg/mL (A), 0.005 mg/mL (B), 10 μM (C), and 10 μM (D), respectively. In this cell line, the combination of 5 μM OL + 10 μM HT demonstrated a synergistic effect in the baseline cells (E). For IM-ConjEpi cells, ROS levels were reduced significantly by CONV, OPT3, HT, and OL at 0.05 mg/mL (F), 0.005 mg/mL (G), 25 μM (H), and 25 μM (I), respectively. Furthermore, 5 μM of OL + 10 μM of HT had an increased activity on UV-B stimulated IM-ConjEpi cells, compared with each compound alone (J). Data are presented as relative fluorescence units (RFU) normalized to micrograms (μg) of total protein for three independent experiments (performed in duplicate) ± SEM. * p < 0.05, ** p < 0.01, *** p < 0.001, compared with vehicle-treated-UV-B stimulated cells; + p < 0.05, ++ p < 0.01, +++ p < 0.001, compared with control cells. Vehicle: Cell culture medium (E,J).
Regarding IM-ConjEpi cells, CONV and OPT3 significantly decreased ROS levels in both UV-B exposed and non-exposed cells, from 0.05 mg/mL (Figure 8F) and 0.005 mg/mL (Figure 8G), respectively. HT and OL also demonstrated a significant antioxidant effect, both from 25 μM, while ROS production was prevented in non-exposed cells from 1 μM for HT (Figure 8H) and 5 μM for OL (Figure 8I).
In terms of OL + HT combination, from low concentrations of 5 μM of OL + 10 μM of HT, the mixture prevented and decreased UV-B-induced intracellular ROS production in both cell lines (Figure 8E,J). The synergistic antioxidant activity can be clearly observed not only in the UV-B exposed IM-ConjEpi cells, but also in the baseline ROS production by HCE cells. The antioxidant effect in IM-ConjEpi non-stimulated cells can be achieved by each compound alone, while the decrease of ROS levels in HCE stimulated cells can be observed by HT alone.
5. Discussion
This work proposes the valorization of an environmentally hazardous agro-industrial by-product into an extract with demonstrated potential as treatment for inflammatory and oxidative stress-related ocular surface diseases. Different extracts derived from OP, together with two of their major phenolic compounds, HT and OL, were tested in vitro on human corneal and conjunctival epithelial cells for their cytotoxicity, anti-inflammatory, and antioxidant effect.
Regarding the results of cell viability assay for both cell lines, CONV and OPT1 can be used up to 0.80 mg/mL, while OPT2 and OPT3 up to 0.40 mg/mL. HT is nontoxic up to 100 μM, OL up to 300 μM, and their mixture OL + HT up to 5 μM of OL + 50 μM of HT. These data agree with studies already published regarding cytotoxicity of the same or similar phenolic compounds on ocular cell lines. Granner et al.
[30], Zhu et al.
[32], and Liu et al.
[33] used safely HT up to 100 μM in retinal pigment epithelial cells, while Zou et al.
[31] proved the antioxidant effect of HT in the same cell line up to 200 μM. Abengózar-Vela et al.
[13] showed that resveratrol is nontoxic up to 300 μM for ocular surface epithelial cells, while for Paladino et al.
[39], resveratrol can be safely used up to 200 μM for retinal epithelial cells. Stoddard et al.
[40] studied the antioxidant activity of four different polyphenols (quercetin, n-propyl gallate, epigallocatechin gallate (EGCG), and gallic acid) in human corneal cells and demonstrated that their toxic concentrations could vary from 41.3 to 337.5 μM, depending on the compound. Regarding cytotoxicity studies of crude extracts on human corneal cells,
Aloe vera extract can be used up to 0.13 mg/mL
[41], Maple leaves extract up to 0.20 mg/mL
[42], and
Polygonum cuspidatum aqueous extract up to 100 mg/mL
[43]. Importantly, as also can be observed from the four different extracts produced from the same material in this study, the extract composition can vary, depending on the extraction method and conditions. This can affect the maximum allowable concentration of each extract.
Our results also showed that, among the four extracts derived from OP tested, CONV (produced by conventional solid–liquid extraction) and OPT3 (generated by sequential scCO
2 de-oiling, followed by PLE optimization) demonstrated a strong anti-inflammatory activity in a dose-dependent manner. Both inhibited TNF-α -stimulated IL-6, IL-8, IL-1β, and IP-10 secretion by ocular surface corneal and conjunctival epithelial cells. HT was also able to decrease the levels of the aforementioned interleukins in both cell types. OL demonstrated significantly less anti-inflammatory effect than HT or OP extracts, inhibiting only IP-10 stimulated production in conjunctival cells. However, when OL is combined with HT, the mixture can inhibit IP-10 secretion by HCE cells at very low concentrations, at which each compound alone would have no significant activity. This fact proves that the compounds can demonstrate a remarkable synergistic anti-inflammatory effect when combined. Increased levels of the measured cytokines/chemokines have been related to several inflammatory ocular surface diseases, such as DES
[44][45][46][47][48][49][50][51][52][53][54] and ocular allergy
[55][56][57][58]. More recent findings reveal altered levels of IL-17A in conjunctiva of DES patients, proving that IL-17A plays an important role in the conjunctival epithelial disruption
[59]. High levels of IL-17A were also detected in mice stimulated by desiccating stress
[46]. However, in our study, TNF-α failed to stimulate IL-17A production in both ocular surface cell lines selected; additionally, in some of our experiments on HCE cells, IL-1β secretion upon TNF-α-stimulation was not sufficient. Another proinflammatory stimulus should be tried in the future either alone or in combination with TNF-α.
In terms of extracts’ composition, as already described in
Table 1, CONV contained 1.7 mg of HT/g of DE and 3.4 mg of OL/g of DE, OPT1: 4.4 mg
HT/g and 5.7 mg
OL/g, OPT2: 2.9 mg
HT/g and 11.4 mg
OL/g, and OPT3: 7.7 mg
HT/g and 0.0 mg
OL/g. Hence, at a concentration of 0.40 mg/mL (used for the first screening of anti-inflammatory activity), 4.4 μM of HT and 2.5 μM of OL were tested for CONV, 11.4 μM of HT and 4.2 of μM of OL for OPT1, 7.5 μM of HT and 8.4 μM of OL for OPT2, and 20.0 μM of HT and 0 μM of OL for OPT3. As already mentioned, HT demonstrated a much stronger inhibition of the secretion of the selected interleukins/chemokines than OL on both cell lines. Therefore, because OPT3 had the strongest anti-inflammatory activity, this can be probably related more to HT (or similar compounds) than OL. OPT2 had the highest concentration of OL; however, it demonstrated very poor inhibition of the selected cytokine/chemokines. This fact can also support this hypothesis. On the other hand, OPT3 comprises many different phenolic compounds apart from HT. As can be observed from
Table 1, OPT3 is the extract with the highest total phenolic content (2.6 times more than the CONV) and simple phenols’ concentration (like HT)
[38]. For OPT3, 0.200 mg/mL comprise 10.0 μM HT and 0 μM OL, while 0.050 mg/mL contains 2.5 μM HT and 0 μM OL. For CONV, 0.20 mg/mL comprises 2.2 μM HT and 1.3 μM OL, 0.40 mg/mL includes 4.4 μM of HT and 2.5 μM of OL, while 0.60 mg/mL contains 6.6 μM of HT and 3.8 μM of OL. Comparing the effective concentrations of OL and HT alone with those included in the concentrations of the extracts tested, a clear synergistic effect can be observed in the extracts. This fact has already been proven by the combination of OL + HT. All these data can explain why OPT3 demonstrate a stronger anti-inflammatory activity compared with CONV, confirming our hypothesis.
In addition, all extracts and compounds tested demonstrated a significant antioxidant activity in a dose-dependent way, starting from very low concentrations, inhibiting UV-B induced ROS production in corneal and conjunctival cells. The combination of OL + HT demonstrated a strong synergistic antioxidant effect in both cell lines either in the stimulated (for IM-ConjEpi) or in the baseline (for HCE) cells. ROS are produced during oxidative metabolism as by-products and are formed by partial reduction of oxygen
[60][61]. Oxidative stress is the disruption of the balance between the antioxidant and the pro-oxidant system of the cells, many times leading to oxidative damage
[62]. The oxidative damage has been involved in the pathophysiology of many chronic systematic diseases, like cancer, inflammation, and neuro-degenerative disorders
[2]. Ocular surface epithelial tissues are exposed to atmospheric oxygen and UV rays of the sunlight
[63], also being vulnerable to oxidative stress-induced ocular diseases. Accumulated ROS in tears and Meibomian gland have been related to ocular tissue inflammation and damage
[64]. Oxidative damage has been proven to be involved in the pathophysiology of several ocular surface diseases, such as conjunctivochalasis
[6], atopic keratoconjunctivitis
[7], and Sjogren and non-Sjogren DES
[7][9][65][66]. Further, Birkedal-Hansen et al.
[67] proved that chronic exposure to environmental stress upregulates oxidative biomarkers, causing loss of the regenerative ability of the corneal cells.
At the concentration used during the first screening of antioxidant activity (0.05 mg/mL), 0.6 μM of HT and 0.3 μM of OL were tested for CONV, 0.18 μM of HT and 0.5 of μM OL for OPT1, 0.9 μM of HT and 1.1 μM of OL for OPT2, and 2.5 μM HT and 0 μM OL for OPT3. OPT3 at 0.005 mg/mL comprises 0.3 μM HT and 0 μM OL. CONV at 0.20 mg/mL contains 2.2 μM HT and 1.3 μM OL, while at 0.05 mg/mL, it contains 0.3 μM of HT and 0.6 μM of OL. HT and OL alone can inhibit ROS production by HCE cells at 10 μM and by IM-ConjEpi at 25 μM. From the combination of OL + HT, a synergistic antioxidant effect was observed. As already explained above, both extracts comprise several phenolic compounds and different total phenolic content (
Table 1)
[38]. In addition, from the results of both cell lines, it can be observed that OPT3 demonstrates a strong inhibition of the ROS production in 10 to 40 times lower concentrations compared with CONV. Therefore, the synergistic antioxidant activity of several phenolic compounds present in the extracts can be confirmed.
The beneficial anti-inflammatory and antioxidant effects of other pure phenolic compounds on the ocular surface have already been reported. For instance, previous studies from our group demonstrated that quercetin and resveratrol, either alone or in combination, inhibited IL-6, IL-8, and IP-10 secretion by TNF-α–stimulated human corneal and conjunctival cells
[13]. They also reduced IL-4, IL-1α, IL-2, and TNF-α tear production in mice exposed to desiccating stress and decreased corneal staining when administrated as eye drops
[14]. CD4 T-cell conjunctival infiltration was also decreased in adoptively transferred mice with T cells from desiccating stress-exposed animals treated with quercetin and resveratrol. In addition, both compounds were able to inhibit the UV-B induced oxidative stress in human corneal and conjunctival cells, in a dose-dependent manner
[13]. Further, several studies have demonstrated the in vitro anti-inflammatory activity on human corneal cells of curcumin
[68], DA-6034 (7-carboxymethyloxy-3′4′,5-trimethoxy flavone)
[69], Daidzin
[70], EGCG
[71][72][73][74], Pterostilbene
[75], and a mixture of ferulic acid with kaempferol
[76]. Catechin was also able to reduce Prostaglandin E2 production on rabbit cornea cells
[77]. A strong antioxidant activity on human corneal cells has been reported only for Pterostiblene
[75], Daidzin
[70], and EGCG
[71]. However, there are few studies exploring and establishing the relation between the antioxidant and the anti-inflammatory activity of natural phenolic compounds on human ocular surface cells. In addition, all these studies examined the effect of the molecules only on corneal cells and not on any additional ocular surface cell line. Our work presents an in vitro comparative study for the olive phenolic compounds between two different parts of the ocular surface (conjunctiva and cornea) exclusively of the human epithelium. Thus, it proposes a promising therapy for oxidative- and inflammatory-related diseases of the entire human ocular surface.
The antioxidant activity of the phenolic compounds has been attributed to the presence of the quinone/semiquinone/hydroquinone triad equilibrium, the chain-breaking scavenging effect of aromatic H atom donors, and the hydroxyl substitution of the aromatic ring
[78][79][80]. Regarding the latest, the position and number of the aromatic hydroxyl groups in the structure of the phenolic molecule can affect its final ROS scavenging capacity
[81]. The anti-inflammatory activity of these molecules has also been related to the position of these groups. In our study, HT was the molecule with the highest inhibitory activity of the measured ILs. For HT, the hydroxyl groups have been found to demonstrate strong interaction with the ASP348 protein of sirtuin 1 enzyme, upregulating its expression, and hence downregulating inflammatory response by vascular endothelial cells
[82]. More recently, hydrogen bond interactions have been confirmed between the hydroxyl groups of HT and the Ser-530 residue of cyclooxygenase-2, inhibiting the expression of the enzyme
[83]. It is also important to observe that TY, which does not have the 3-hydroxyl of HT, does not have any affinity with the enzyme, confirming the importance of the position of these groups
[83].
Apart from individual phenolic compounds, numerous crude polyphenolic-rich extracts have also demonstrated strong in vitro antioxidant and/or anti-inflammatory activity on the ocular surface. Preliminary studies on fractions of olive polyphenolic extracts have already demonstrated interesting inhibition of ROS and TNF-α production by UV-B-induced and LPS-induced rabbit epithelial corneal cells, respectively
[37]. However, our work consists of a complete pharmacological study of four different interleukins/chemokines secretion by two different cell lines of the human epithelium not only for olive extracts, but also for their two major phenolic compounds. Further, it compares extracts with different composition obtained from the same material, explaining their distinct activities. Regarding different plant materials, extracts from
Camellia japonica [84],
Chamaecyparis obtuse [85],
Aloe vera [41],
Euphrasia officinalis [86],
Lamium album [87],
Polygonum cuspidatum [43], and Maple leaves
[42] have also been found to reduce inflammatory biomarkers in human corneal cells. Hence, the potential of naturally derived polyphenols and polyphenols-rich extract is becoming an emerging issue in the field.
In summary, our results have demonstrated that a convectional and an HT- enriched optimized extract derived from OP, along with the major OP pure phenolic compounds HT and OL, can inhibit inflammation and oxidative damage in human corneal and conjunctival epithelial cells. However, there are some limitations in this study. Although TNF-α acted good as inflammatory stimulus, different inducers could be used to achieve the stimulation and measurement of IL-17A, TNF-α itself, and/or any other interleukin/chemokine not stimulated by TNF-α. Further, although HCE has been proven to demonstrate high correspondence to the human tissues of origin
[88], IM-ConjEpi is a relatively new commercial SV-40 Large T antigen immortalized cell line from primary human conjunctival epithelial. However, according to the manufacturer, it has a 99% purity and maintains expression of epithelial specific markers such as CK18 and CK19. Finally, it is also true that in vivo models are much more complex. For many of the compounds
[69][70][73][74][76][89][90][91] and extracts
[43][84][85][92][93][94][95] mentioned in the bibliography, the anti-inflammatory activity has also been proven in vivo. For EGCG
[96] and Daidzin
[70], the in vivo antioxidant effect has also been studied. Hence, in vivo studies in a desiccating stress mice model are currently being performed for selected extracts and compounds, to support our in vitro findings.
In conclusion, extracts derived from OP and their major phenolic compounds, particularly HT, seem to be promising potential treatment for ocular surface inflammatory diseases, such as DES. The use of these type of OP extracts is of chief importance for the green development of related industries, as they propose a high value application of an agro-industrial by-product. The results of this study illustrate how the sustainable and intensified extraction techniques are proved to be competitive compared to the conventional ones, while a high selectivity towards biomarkers related to dry eye is established.
 +1 point
+1 point