1000/1000
Hot
Most Recent
 +1 point
+1 point
The pantropical genus Vanilla is a member of the Orchidaceae family. It originated in America and differentiated in America, Africa, and Asia. About 132 species of Vanilla have now been accepted since the discovery of the genus. Within the Vanilla genus, leaflessness has appeared at least three independent times, in Africa, Asia, and America as an adaptation to drought conditions. The South-West Indian Ocean (SWIO) region is home to seven leafless species. Chloroplast markers failed to differentiate species within this recent SWIO clade. Morphological traits used for description are very similar between species and do not provide reliable identification. Leafless Vanilla species from the SWIO islands are thus an excellent model for understanding the evolution of orchids. For their conservation, and to confirm the identity of these leafless species, an integrated approach with classical taxonomy using a large number of samples, intense fieldwork on biology and ecology, and molecular studies using variable markers is necessary.
With more than 24,000 species and 700 genera worldwide, Orchidaceae is one of the largest families of flowering plants [1][2], and originated around 83 Mya (Million years ago) [3][4]. Orchids are widely celebrated for their beautiful flowers, extraordinary diversity, and potential medicinal proprieties. For example, among the 494 terrestrial and epiphytic orchid species recorded in Southern Africa, 49 are used as traditional medicinal plants to treat a wide range of diseases [5]. The South-West Indian Ocean (SWIO) islands are one of the global hotspots for orchids. The Orchidaceae family is represented by 65 genera and over 1000 species in this region [6]. Madagascar is the richest, with approximately 862 species, of which more than 85% are endemics [7], and 57 genera [8]. The Orchidaceae genus Vanilla can be considered as a flagship genus in the SWIO region, because the genus is represented there by wild, as well as cultivated, genetic resources. This region ranks first in terms of leafless Vanilla species richness [9][10]. It is also renown worldwide for its Vanilla fruit production, with the islands of Madagascar, Comoros and La Réunion being amongst the first world producers [11].
Vanilla plants are perennial vines characterized by thick and fleshy stems [12], a monopodial growth habit, aerial roots growing at each node, absence of pseudobulbs, fleshy fruits and wingless seeds [9]. The inflorescence is axillary [13]. The aerial roots are hairless, while the underground roots are hairy [14][15]. Leafless Vanilla species share most vegetative characteristics with leafy species, but they are photosynthetic plants with green, sometimes reddish to glaucous stems, distinguished by the absence of leaves or the modification of the leaves into scale-like structures [10][16]. Also, as opposed to leafy species, green stems of leafless species do not have a sclerenchyma band separating cortex from ground tissue [14]. Two types of roots were identified in leafless species from Madagascar: short, flattened or filiform roots used for attachment to a supporting tree, and long aerial roots up to 15 m exploring the soil for nutrition [15]. The same study demonstrated that populations in the north, where rainfall is higher, have thinner aerial roots with smaller xylem vessels than those in the arid south [15]. This is probably one of the developed strategy of succulent plants to face the scarcity or abundance of water in the soil [17]. Floral morphology is diverse but, in general, flowers are composed of three sepals, two petals, and a third large petal called labellum [10][12] forming a funnel surrounding the column [13]. Pollinia and stigma are separated by a membrane called the “rostellum”. It usually prevents self-pollination within the same flower, although species are self-compatible [18][19]. The number of opened flowers per day varies from one to several, depending on the species. For Mexican leafy species, for example, it is generally one (e.g., V. phaeantha) to three (e.g., V. calyculata) [20]. In Madagascar, we observed that the number of opened flowers per inflorescence per day can go up to three in the leafless white-flower species (V. madagascariensis, V. decaryana, V. bosseri), whereas this number is on average one flower and rarely two in the yellow-flower populations of Montagne des Français (V. humblotii). This is also the case of V. humblotii from Mayotte, for which on average 1.84 flowers open per day and per individual [21]. Flowers usually open in succession and are short lived (one day) (e.g., in leafless V. barbellata, V. claviculata, V. dilloniana [22]) but can remain for two or three days for some species like the leafy species V. inodora [20]. Unfertilized flowers rapidly fall, otherwise they turn into fruits in the form of an elongated capsule, called a “pod”.
The genus Vanilla Plum. ex Miller forms a monophyletic group (34.6 Mya) (Figure 1) in the large Orchidaceae family, the Vanilloidae sub-family, the Vanilleae tribe and the Vanillinae sub-tribe [1][10][16][23].
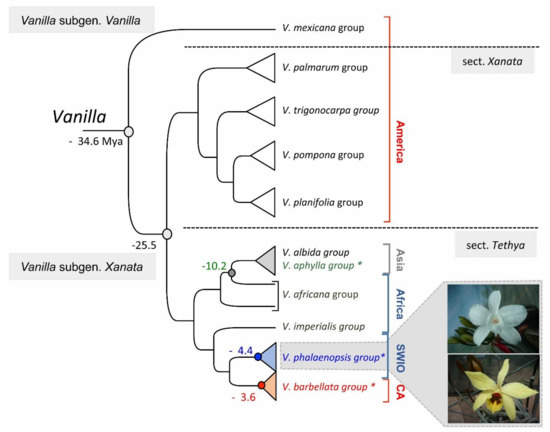
Figure 1. Representation of the (rbcL, psaB, psbB, psbC) phylogeny of Bouetard et al. in 2010 including species groups defined by Soto Arenas and Cribb in 2010, showing leafless Vanilla species names in color (green for Asia, blue for SWIO, red for Caribbean CA) [3][9]. Node dates in Mya correspond to datations obtained by Bouetard et al., with the datation of Vanilla node at −34.6 Mya estimated using 71 Mya as the minimum age for Vanilloideae published by Ramírez et al., 2007, based on fossil evidence of an orchid pollinarium (of Meliorchis caribea) attached to an extinct stingless bee (Problebeia dominica) recovered in Miocene amber [3][4].
Vanilla is a transoceanic genus, found only in tropical forests between the 27th north and south parallels, but absent in Australia (Figure 2).
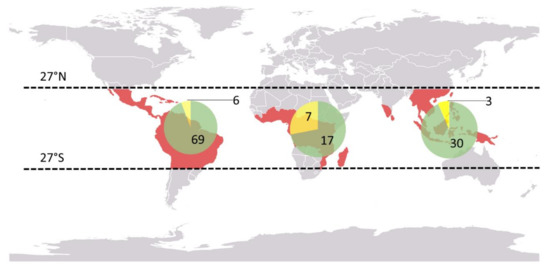
Figure 2. World distribution of Vanilla species: number of leafy Vanilla species (in green) and number of leafless species (in yellow) based on the World checklist of Selected Plant Families [24], the International Plant Name Index [25] and the database of Tropicos [26].
According to Rolfe, the diversification of the genus happened in Brazil and Guiana [23]. However, based on floral morphology, Portères suggested a primary diversification center in Indo-Malaysia that has evolved into two groups (Madagascar, Mascarene Islands, Africa versus Asia and Western Pacific islands), with the American species resulting from the migration of Asian species during the Tertiary (65.5–2.5 Mya) [10]. But the recent molecular phylogeny rejected this theory of Portères and suggested a South American center of diversity [3]. In 2005, Cameron stated that the differentiation of the genus Vanilla would have taken place before the dissolution of Gondwana [27], about 129 Mya [3]. However, following a fossil-based tree dating in Orchids [4], Bouetard et al. supported the alternative theory that Vanilla genus probably appeared 34 Mya (Figure 1), and its migration from the New World to the Old World occurred at the beginning of the Tertiary, about 25 Mya via a transoceanic dispersion, facilitated by the proximity of the continents and the presence of the series of islands of the mid-oceanic ridge of the Rio [3]. Three obvious cases of transoceanic migration events from Africa were also noted: (1) the dispersal of the genus from Africa to Asia; (2) from Africa to the SWIO islands and finally (3) from Africa to the Caribbean islands [3], the two last events concerning therefore the evolution of leafless groups in the SWIO and Caribbean islands.
Vanilla orchids are thermophilous [14]. These terrestrial, epiphytic or hemi-epiphytic plants are adapted to a variety of habitats [10][14][16][20] although they are scarce on mountain rain forest [14]. Lamellosae are found in fresh and wet places. They are mainly ombrophilous to mesophilous with various levels of heliophilia [10]. Papillosae species are also sciaphilous, living in rainforest formations, to heliophilous, sometimes growing in tropophilous and sub-xerophilous formations [10]. V. organensis, V. palmarum, V. wrightii and relatives without lamellae and with column fused to the labellum can be found in more mesophilic or even tropophilic formations in the area of Brazilian palm groves or wooded savannas [10]. Species with longitudinal lamellae and aromatic fruits are distributed in coastal or sublittoral forests [10]. However, some can be found in uncommon habitats like marshy forests (e.g., V. arcuata and V. paludosa [28]). While most species can climb to various substrates (trees and rocks), an epiphytic Vanilla (V. palmarum) is exclusively associated with nine species of palms in South America [29]. Some species can be found in different types of ecosystems like V. inornata [30]. There are species that thrive on particular substrates such as V. martinezii growing on an unusual peat-like soil [20], V. planifolia alliance in Central America restricted to calcareous or sedimentary substrate [14] and V. insignis found on seasonally dry volcanic areas [14]. Vanilla species are often associated with mycorrhizal fungi [15][16][31]. For example, studies in Puerto Rico, Cuba and Costa Rica recorded several species belonging to three genera of mycorrhizal fungi (Ceratobasidium, Thanatephorus and Tulasnella) in V. potaei and V. planifolia [31].
Vanilla orchids are thermophilous [14]. These terrestrial, epiphytic or hemi-epiphytic plants are adapted to a variety of habitats [10][14][16][20], although they are scarce on mountain rain forest [14]. Lamellosae are found in fresh and wet places. They are mainly ombrophilous to mesophilous with various levels of heliophilia [10]. Papillosae species are also sciaphilous, living in rainforest formations, to heliophilous, sometimes growing in tropophilous and sub-xerophilous formations [10]. V. organensis, V. palmarum, V. wrightii and relatives without lamellae and with column fused to the labellum can be found in more mesophilic or even tropophilic formations in the area of Brazilian palm groves or wooded savannas [10]. Species with longitudinal lamellae and aromatic fruits are distributed in coastal or sublittoral forests [10]. However, some can be found in uncommon habitats like marshy forests (e.g., V. arcuata and V. paludosa [28]). While most species can climb to various substrates (trees and rocks), an epiphytic Vanilla (V. palmarum) is exclusively associated with nine species of palms in South America [29]. Some species can be found in different types of ecosystems like V. inornata [30]. There are species that thrive on particular substrates such as V. martinezii growing on an unusual peat-like soil [20], V. planifolia alliance in Central America restricted to calcareous or sedimentary substrate [14] and V. insignis found on seasonally dry volcanic areas [14]. Vanilla species are often associated with mycorrhizal fungi [15][16][31]. For example, studies in Puerto Rico, Cuba and Costa Rica recorded several species belonging to three genera of mycorrhizal fungi (Ceratobasidium, Thanatephorus and Tulasnella) in V. potaei and V. planifolia [31].
Aphyllae is the only group that contains species adapted to hot, xeric, full sun habitats [10][14][16]. They grow on coastal bushes (e.g., V. roscheri and V. decaryana), in very dry tropophilous forests (e.g., V. madagascariensis), on sandy soils (e.g., V. perrieri, V. bosseri, V. aphylla), on rocks (e.g., V. humblotii, V. phalaenopsis and V. marmoreisense), on coral reefs (V. roscheri) but also in mangrove swamps (e.g., V. barbellata and V. dilloniana) or in humid forests (V. madagascariensis) (Figure 3A,B) [8][10][32][33][34].

Figure 3. Ecology and usages of leafless Vanilla species: (A) V. phalaenopsis individual growing on a rock in Seychelles, (B) V. decaryana plant in thorny bushes in the south of Madagascar (Antandroka) and (C) leafless Vanilla stems for sale on the Tuléar market (Madagascar). (Photographies A, C: Pascale Besse, B: Cathucia F. Andriamihaja).
Leafless species are exclusively found on islands or on nearby littoral environment [10]. According to our observations in several populations in Madagascar, they are all hemi-epiphytes characterized by early germination and early stage in the soil, a root system exploring the ground and a long stem climbing along a support.
In most cases, Vanilla species combine vegetative and sexual reproduction, except for V. mexicana, V. bicolor and V. palmarum for which artificial clonal propagation was reported to be unsuccessful or difficult [18][35][36]. The vegetative reproduction is possible as soon as an individual or a sufficient fragment of stem cutting can take root in the soil [35], sprout vegetative buds [36] and climb to a support to create a new individual. This clonal propagation constitutes the predominant reproductive, colonization and survival strategy. Studies on leafless species also revealed their ability for clonal reproduction in natural habitats. In Puerto Rico, Nielsen and Siegismund used polymorphic enzymes to analyse genetic diversity [22][37]. The results showed a high clonality rate for V. dilloniana (38%) [22], a moderate rate for V. barbellata (25%) [22] and a low rate for V. claviculata (6%) [37]. In V. humblotii in Mayotte, microsatellite analyses showed a moderate clonality rate (12.5%), with an average clonal patch size of 4.6 (±2.7) m, and a phalanx distribution [21].
The sexual reproduction, on the other hand, can occur only during the flowering period and only when the individual has attained maturity [20]. Also, it is common that some populations of a species fail to bloom for one or more successive years, as it was reported for V. hartii in tropical America [20]. There are, however, species that bloom throughout the year or during several months like V. inornata [30], V. rivasii [38] or V. bicolor [36]. Although some Vanilla species have an unusual high natural fruit set (>40%) (Figure 4), which suggests self-pollination [20][36][39][40], most are pollinator-dependent due to the presence of the rostellum membrane separating the female and male parts of the flower [10][19][21][23]. Some mechanisms were reviewed by Gigant et al., to explain self-pollination: it may be caused by either stigmatic leak and/or the presence of a dehydrated or reduced rostellum, or agamospermy (e.g., V. bicolor and V. palmarum), but some spontaneous self-pollinations remain unexplained (e.g., V. planifolia and V. chamissonis) [18]. In addition to these mechanisms, Householder et al. also noted the absence of a rostellum in V. guianensis in Peru, facilitating pollen transfer between the two sexual organs [36].
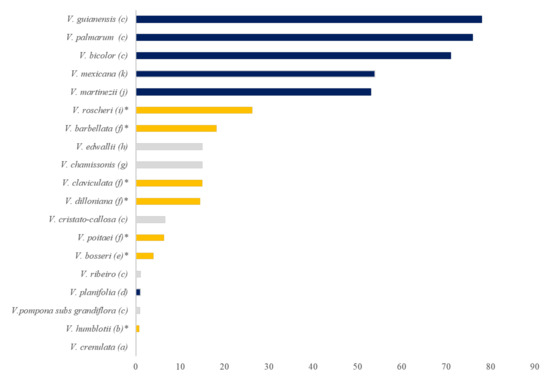
Figure 4. Rate of natural fruit set (%) of Vanilla species according to (a) Soto Arenas and Cameron [14], (b) Gigant et al. [21], (c) Householder et al. [36], (d) Ackerman et al. [41], (e) Petersson [42], (f) Tremblay et al. [39], (g) Macedo [43], (h) Pansarin, Aguiar, and Pansarin, 2014, (i) Gigant et al., 2014, (j) Soto Arenas and Dressler, 2010, (k) Gigant et al. [40]. Species that can self-pollinate spontaneously are in dark blue, leafless Vanilla species are in orange and indicated by *, and the grey bars represent the other species.
Pollinators visit flowers for various reasons, such as collecting nectar, oil, pollen, stigmatic exudates or floral scents [44][45][46]. In Vanilla, floral odor seems to play an important role in attracting pollinators as in the case of V. trigonocarpa or V. pompona in America [14][20][36]. Other strategies are also used by orchid species to attract pollinators such as deceptive systems [47], as described for V. planifolia, V. odorata, and V. insignis in America [18]. In orchids, bee-pollination also requires a morphological association between floral characteristics and the insect traits [44][48][49][42].





