Alzheimer’s disease (AD) is a neurodegenerative disease with a high incidence in the elderly. Many preclinical studies show that a natural product, ferulic acid (FA), displays neuroprotective effects in AD models.
1. Introduction
Alzheimer’s disease (AD) is an age-related progressive degenerative disease of the central nervous system with multiple causes. The onset of AD is closely related to age, genetic factors, and environmental factors. However, None of any existing theories can fully explain the complex pathology of AD at present. The clinical manifestations are memory and cognitive function impairment usually accompanied by mental behavior abnormality and personality change
[1]. Its characteristic pathological feature is the deposition of abnormal protein aggregates, namely β amyloid protein (Aβ) in extraneuronal space (senile plaques) and hyperphosphorylated tau protein within neurons (neurofibrillary tangles).
AD is distributed worldwide and is the most common cause of dementia. With the gradual aging of the population in developed and developing countries, AD has become the fourth leading cause of human death and has a huge economic impact
[2]. It is estimated that, by 2050, more than 1.315 billion people worldwide will have AD
[3].
Ferulic acid (FA), 4-hydroxy-3-methoxycinnamic acid (C
10H
10O
4, MW = 194.18), is a type of phenolic acid and a derivative of cinnamic acid. It is widely present in nature and distributed in a variety of fruits, vegetables, and grains, and also one of the active components in some Chinese medicinal herbs such as
Angelica acutiloba,
Ligusticum striatum,
Cimicifuga dahurica, and
Crocus sativus [4][5]. A variety of herbal medicines containing FA are believed to have the effects of promoting blood circulation, removing blood stasis, and reducing inflammation and pain in traditional Chinese medicine. Modern medical research has found that FA is a multi-functional compound with anti-inflammatory, antioxidant, anti-amyloid, neurotrophic, anti-microbial, antiallergic, hepatoprotective, anti-carcinogenic, and anti-thrombotic activities
[6][7][8]. Recently, many studies have shown that FA is neuroprotective in a variety of neurodegenerative diseases models, both in vivo to
in vitro, and especially in AD models.
2. Ferulic Acid in Animal Models of Alzheimer’s Disease
Animal experiments are indispensable for human health science research. In recent years, with the rapid development of biotechnology, the requirements for animal welfare and animal ethics have gradually strengthened. The 3R principle (replacement, reduction, refinement) is one of the representative products. Using systematic reviews to conduct targeted statistical analysis of published animal-related experiments can help researchers make full use of existing data, avoid unnecessary repetitive experiments, optimize existing experimental designs, and reduce cost.
2.1. Mechanism of FA in Anti-AD
The pathogenesis of AD is complex, and the etiology has not been fully elucidated. The current hypothesis involves many aspects such as Aβ toxicity, tau hyperphosphorylation, neuroinflammation, oxidative stress, and abnormal immune function.
Reducing amyloid
[9][10][11][12][13], reducing inflammation
[9][14][11][12], antioxidant activity
[15][10][16][11][17][12], repairing mitochondrial damage
[18][13], and inhibiting microglial
[19][20][21] or astrocyte activation
[22][23] are the main mechanisms (
Figure 1):
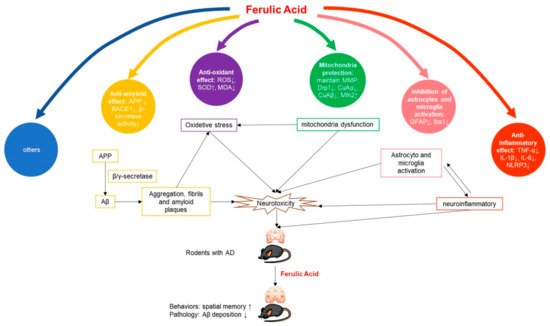
Figure 1. Potential mechanisms of FA in AD animal model.
2.1.1. Anti-Amyloid Effect
The deposition of amyloid plaques, especially Aβ, is the most prominent pathological feature in the development of AD.
In vitro aggregation tests have shown that FA can inhibit the formation and extension of Aβ by affecting the fiber elongation process
[24]. This effect has been verified on the genetically modified
Caenorhabditis elegans model
[25].
In the body, Aβ is produced by β-secretase and γ-secretase through continuous hydrolysis of amyloid precursor protein (APP). Among them, β-secretase is considered to be the rate-limiting enzyme for Aβ production
[26]. Research by Ji-jing Yan et al.
[9] showed that FA inhibited Aβ deposition in the cortex, but beneficial effects were observed only in the low-dose group (5.3 mg/kg/d). Beta-site APP cleaving enzyme 1 (BACE1) is a major β-secretase. Takashi Mori et al.
[10][11][12] reported that FA can reduce amyloid APP metabolism by reducing BACE1 expression and β-secretase activity in mice, thereby improving Aβ
1-40 and Aβ
1-42 deposition in the brain parenchyma and cerebral vessels of AD mice and slowing related toxic reactions. The team confirmed this result with
in vitro experiments. FA not only breaks down pre-formed fibrous tangles, but also inhibits the formation and extension of Aβ in a dose-dependent manner
[12]. Wang Qian et al.
[13] found that FA can effectively reduce the expression levels of APP, BACE1, total Tau, and Tau pS396 in the brains of AD model mice, and increase the expression of PSD95 and PD-HE1α. The above results indicate that FA may have a therapeutic effect on AD by directly inhibiting the formation of amyloid plaques.
2.1.2. Anti-Inflammatory Effect
In recent years, more and more studies have revealed that inflammation plays an important role in the occurrence and development of AD. The anti-inflammatory effects of FA have been reported. Takashi Mori et al.
[10][11][12] have shown in multiple studies that FA alleviated neuroinflammation in PS1/APP mice, including Aβ plaque-related proinflammatory cytokines TNF-α and IL- 1β expression. Ji-jing Yan et al.
[9] also found that FA reduced the level of IL-1β in mouse brain cortex. Ming Rui et al.
[21] found that FA can reduce the increased expression of IL-1β, IL-6 and TNF-α in the brain tissue of AD model mice. Huang Hao et al.
[14] found that FA treatment can alleviate the LPS-induced increase in IL-1β, caspase-1, NLRP3, and PDE4B mRNA levels, which means that FA has the potential to block the activity of NLRP3 inflammasome.
2.1.3. Antioxidant Effect
The hypothesis that oxidative stress is involved in AD development has been confirmed by multiple laboratories
in vitro and
in vivo [27][28][29]. A review by Butterfield
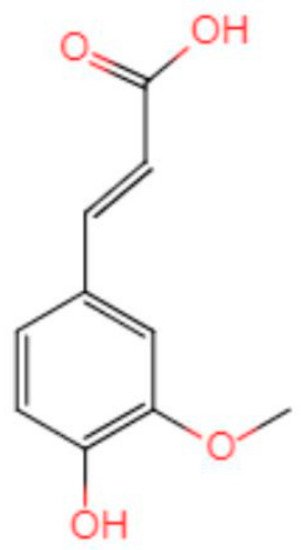
[30] shows that numerous studies have found that Aβ produces neurotoxicity by inducing oxidative stress (OS) in the brain. The chemical structure of FA determines its antioxidant function (
Figure 2). The presence of 3-methoxy and 4-hydroxy groups on the benzene ring can easily form resonance-stable phenoxy groups. A carboxylic acid group having an adjacent unsaturated C-C double bond can stabilize the free radical by preventing the free radical film attack. At the same time, the carboxylic acid group can act as a lipid anchor, providing protection against lipid peroxidation
[31]. An RCT study showed that FA supplementation can improve lipid metabolism in patients with hyperlipidemia, and improve oxidative stress and inflammation
[32]. These results suggest that FA may help to improve the oxidative stress state and inflammation in the brain of AD patients.
Figure 2. Chemical structure of ferulic acid.
ROS is a common byproduct of electron leakage from the mitochondrial inner membrane during mitochondrial oxidative phosphorylation. Under normal conditions, ROS is rapidly eliminated by enzymes, but when mitochondria are disturbed, ROS production may exceed the cell’s ability to neutralize it, resulting in oxidative damage
[33]. Mohd Faraz Zafeer et al.
[18] found that long-term oral administration of FA can significantly alleviate the exacerbation of ROS in AD mice models.
Mamiya et al.
[15] found that the increased expression of carbonyl protein, one of the markers of protein oxidative damage, can be inhibited by FA pretreatment in AD animal models, indicating that FA can inhibit OS in the brain of AD animal models. Huang Hao
[14] found that SOD expression in the brain of FA-treated AD mice increased. Takashi Mori et al.
[10][11] showed that mRNA and protein expression of three oxidative stress markers (SOD1, catalase, and GPX1) were reduced in FA-treated PSAPP mice. Fan-Shiu Tsai et al.
[16] found that administration of FA to rats increased SOD activity; attenuated Aβ
1-40-induced activity of Mn-SOD and Cu, as well as Zn-SOD in the cortex and hippocampus; and improved the inhibition of GSH activity in the cortex. Wang Yue et al.
[17] found that, after treatment with FA, AD model mice had increased SOD activity and reduced MDA content, suggesting that FA can reduce ROS in the brain of AD mice.
2.1.4. Mitochondria Protection
Autopsy has revealed abnormal hippocampal mitochondria in patients with AD
[34]. In the cell, mitochondria function in a steady state of division-fusion to meet the energy needs of the body
[35]. Drp-1 is a key effector of mitochondrial fission localized at the outer mitochondrial membrane, and Mfn2 is a marker of mitochondrial fusion. The dynamic balance between the two proteins is essential for maintaining the mitochondrial health of neurons
[36].
Mohd Faraz Zafeer et al.
[18] found that FA treatment can restore the balance between mitochondrial division and fusion by regulating the level of PGC-1alpha protein, thereby preventing the loss of mitochondrial membrane potential and reducing Drp-1-dependent mitochondrial fission. Wang Qian et al.
[13] found that the expression of Drp1, CnAα, and CnAβ mRNA and protein in the cerebral cortex of AD mice were significantly reduced in the FA-treated group, and the expression of MFN2 protein was increased. The data suggested that FA can reverse the Aβ-induced, abnormally increased expression of Drp1 and its phosphorylation regulatory pathway genes CnAα and CnAβ. In addition, FA may enhance the resistance of neurons to Aβ toxicity by locating at mitochondria where they can repair mitochondrial biological transport balance, thereby maintaining neuronal signaling delivery and improving learning and cognition in AD mice.
2.1.5. Inhibition of Astrocytes and Microglia Activation
Astrocytes are the most widely distributed and the largest type of glial cells in the brain; they have many functions such as support and protection neuron function, promote neuron repair, and improve neurotransmitter regulation. Glial fibrillary acidic protein (GFAP) is a marker of astrocyte activation. When neurons are damaged, astrocytes are activated to produce a large amount of GFAP. Continuous injection of rats with Aβ
1-40 increases GFAP immunoreactivity
[37]. Mohd Faraz Zafeer et al.
[18] found that long-term oral FA treatment can reduce the expression of GFAP. Jing Beibei et al.
[20] found that, compared with the vehicle group, the level of GFAP decreased with the increase in FA concentration. Morphologically, Ming Rui et al.
[21] observed that, compared with the sham operation group, the number of GFAP-positive astrocytes in the cortex of the model group was significantly increased. After FA treatment, the amount of GFAP positive astrocytes and fluorescence intensity gradually decreased, the axons and dendritic structures of surrounding neurons were basically normal, and the connections between neurons increased significantly. Ji-Jing Yan et al.
[22] confirmed that inhibition of astrocytes by long-term treatment with FA was not mediated by affecting the secretion of neurotrophic factors such as NGF, BDNF, and bFGF. Jae-Young Cho et al.
[23] further found that eNOS, 3-NT, and GFAP colocalized in the brain of AD mice model, and that the change pattern was similar to that of GFAP, suggesting that the ability of FA to protect the brain from Aβ-induced toxicity may be mediated by inhibiting the expression of eNOS and 3-NT in astrocytes. Takashi Mori
[10][11][12] demonstrated that FA treatment can reduce GFAP and Iba1 levels, and inhibit plaque-associated proliferation of microglia and astrocytes. Hee-Sung KIM et al.
[19] suggested that FA induced inhibition of astrocyte activation may be achieved by inhibiting microglial activation. However, the researcher did not provide sufficient evidence to prove this hypothesis.
It is worth noting that Ji-Jing Yan et al.
[22] found that, on the fourteenth day, FA-treated animals showed transient activation of astrocytes manifested by increased expression of GFAP. At the same time, experimental animals performed poorly in behavior with elevated IL-1β. Although the morphology of astrocytes in the hippocampus of mice seemed to return to normal thereafter, they continued to show a weak inhibitory response to Aβ stimulation. This may be the resistance of astrocytes to Aβ toxicity caused by FA treatment. If this is indeed the mechanism by which FA inhibits astrocyte activation, then researchers should further consider whether this resistance can persist for a long time. These data will be helpful to determine frequency and duration of FA medications for AD patients in the future.
2.1.6. Others
Fan-Shiu Tsai et al.
[16] suggested that the mechanism by which FA inhibits Aβ
1-40- induced behavioral injury in rats may be related to the inhibition of AChE activity. Wang Yue et al.
[17] found that the phosphorylated apoptosis-related proteins were significantly increased in the brain of APP/PS1 transgenic mice, while those proteins were significantly reduced in the FA treatment group, indicating that FA may protect neurons by reducing the phosphorylation of apoptosis-related proteins in the brain of AD mice. Mohd Faraz Zafeer et al.
[18] found that long-term oral administration of FA can reduce caspase 3 activation. Huang Hao et al.
[14] suggest that FA may have a significant regulatory effect on the PDE4/cAMP/CREB signaling pathway.
2.2. Anti-AD Potential of FA
The clinical symptoms of AD are mainly progressive cognitive dysfunction and memory impairment, while the characteristic pathological changes are the formation of neuronal fibrillary tangles and senile plaques in the brain.
Compared with vehicle, FA treatment could improve the memory impairment and decrease Aβ deposition in the brains of AD animal models. It is worth noting that, although most studies have shown that Aβ deposition is related to animal behavioral performance, some studies have shown that there is no good correlation between Aβ deposition in the brain and behavioral disorders in transgenic mice expressing APP mutations
[38]. Other studies have suggested that oligomeric Aβ may be the substance that causes AD neurotoxicity
[39][40]. Therefore, we suggest that, when studying the therapeutic effect of drugs on AD models, researchers should choose multiple Aβ-related detection assays, especially the oligomeric Aβ detection assay.
In addition, although we divided the data into different subgroups for analysis based on the species of the experimental animals and whether they were genetically modified, the heterogeneity between the experiments was still large. These differences may be attributed to the different strains of mice used in the experiments. Mice of different strains perform differently. For example, the performance of male C57BL/6 mice is significantly better than that of male Kunming mice, and BALB/c mice are not suitable for the experiments due to fear and poor physical performance
[41][42]. Thus, choosing the right animal strain is one of the prerequisites for successful experiments when conducting such research.
2.3. Development Perspective
2.3.1. Mechanism Exploration
FA has been proven to be effective in treating AD to a certain extent, from cell to animal models; however, the mechanism has not been fully elucidated. Understanding the pharmacological mechanism will greatly facilitate drug development and minimize biological safety concerns for the clinical application of FA. Therefore, there is an urgent need for determining the molecular mechanism of FA for AD treatment.
2.3.2. Structural Modification
Compared with other, larger phenolic compounds, FA has better cell permeability due to its lower molecular weight (194.18 g/mol). Studies have shown that FA has good bioavailability in rats. FA can be quickly absorbed through the gastric mucosa when orally administered, then transported to the portal vein of the liver, combined with glucuronic acid and/or sulfate, and can be maintained in the body for a long time
[43]. However, FA may have difficulty penetrating the blood–brain barrier due to its hydroxyl group. Therefore, structural modification of FA to generate candidate drugs with more potent neuroprotective activity and higher blood–brain barrier penetration is an important direction for future research.
2.3.3. Extrapolation to Humans
In the process of drug development, the ultimate goal of animal experiments is to make the drugs better used in humans. When analyzing the data extracted from the included articles, we found that there is a large heterogeneity between the reported data sets. In addition to laboratory conditions and the genetic background of the animal, the heterogeneity may also be partially derived from differences in administration of the FA. These heterogeneities between experiments can be computer-simulated to establish a physiologically based pharmacokinetic/pharmacodynamic (PBPK/PD) model of drug-to-disease and can use computational pharmacology to improve data integration between different studies for FA on AD animal models. In addition, the establishment of a PBPK/PD model of FA to AD is also conducive to the extrapolation of animal experiments to human experiments, and can help researchers find better routes and dose.
On the other hand, it is worth noting that many natural compounds have been proved to have hormetic dose–response, that is, the compounds have two-way effects on anti-oxidation and anti-neurodegenerative diseases at very low doses and higher doses
[44]. Therefore, it is necessary to carefully select the targeted concentration when extrapolating to the patient. Based on this, more detailed pharmacokinetic studies in animals and humans are needed to determine the effective plasma drug concentration to obtain better and more stable drug effects.
In summary, the current researches of FA on AD are still at the basic research stage. There is no clinic trail being registered on the
clinicaltrials.gov website. It is a long way from preclinical studies on animals to clinical trials on humans, and ultimately, to market. Nevertheless, the results of this review, showing that FA may be effective in repairing the memory and pathological damage caused by AD, suggest that the way is worth travelling.
 +1 point
+1 point