2. Discussion
Salmonella can be transmitted horizontally to chickens from contaminated environmental vectors and vertically from infected hens to offspring. In this study, 1-d-old female AA female chicks, with
Salmonella infection excluded by cloacal swab testing, were reared and challenged under the same conditions, therefore eliminating the influence of genetic background and environment on the experimental results to the greatest extent, and ensured that all the phenotypic results obtained in this study were due to individual differences.
S. enteritidis mainly colonizes the liver, spleen, and intestine of poultry after infection
[25][26], leading to intestinal damage, a decline in growth performance, and even death. Growth performance, pathological changes in organs,
Salmonella loads, and intestinal morphology are important indicators of the severity of
S. enteritidis infection. In this study, compared with
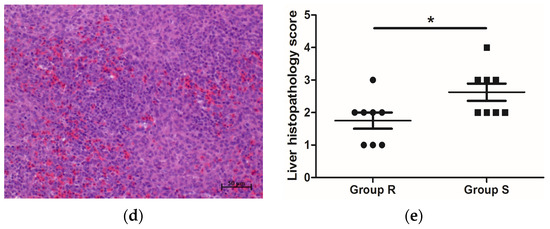
S. enteritidis-resistant chicks, the livers of
S. enteritidis-susceptible chicks became swollen (
Table 3) and displayed salient lesions (
Figure 1b). In addition,
Salmonella loads in the liver and spleen of
S. enteritidis-susceptible chicks were significantly higher than those of
S. enteritidis-resistant chicks (
Table 1). The VCR showed an increasing trend in chicks of group R than in chicks of group S (
Table 2). These results indicated that our grouping scheme, i.e., selecting chicks with differential
S. enteritidis susceptibility, was appropriate, and confirmed the heterogeneous nature of the response of the birds to
S. enteritidis infection.
Figure 1. Histopathology of S. enteritidis-resistant and S. enteritidis-susceptible chicks. (a) Representative liver histopathology of S. enteritidis-resistant chicks (HE staining); (b) Representative liver histopathology of S. enteritidis-susceptible chicks (HE staining); (c) Representative spleen histopathology of S. enteritidis-resistant chicks (HE staining); (d) Representative spleen histopathology of S. enteritidis-susceptible chicks (HE staining). Original magnification, ×200. Black arrows indicate the lymphocytes, yellow arrows indicate the heterophilic cells, and red arrows indicate the lymphocyte nodules in liver tissue. Scale bar = 50 μm. (e) Liver histopathology score of S. enteritidis-resistant and -susceptible chicks, n = 8 per group, Result of significance test was p < 0.05 when marked *.
Table 1. Body weight, tissue index 1, and Salmonella loads of S. enteritidis-susceptible and -resistant chicks 2.
| Items |
Group S 3 |
Group R 3 |
p-Value |
| BW (g) |
229.40 ± 7.47 |
230.88 ± 5.47 |
0.875 |
| Liver Index (%) |
0.043 ± 0.001 a |
0.038 ± 0.001 b |
0.006 |
| Spleen Index (%) |
0.024 ± 0.002 |
0.020 ± 0.001 |
0.158 |
| Liver Salmonella loads (log10CFU/g) |
2.750 ± 0.405 a |
1.152 ± 0.435 b |
0.018 |
| Spleen Salmonella loads (log10CFU/g) |
4.784 ± 0.100 a |
2.491 ± 0.055 b |
<0.001 |
1 Tissue index: Percent of tissue weight relative to body weight. 2 Results are expressed as means ± SEM, with n = 8 per group.3 Group S = selected S. enteritidis-susceptible chicks; Group R = selected S. enteritidis-resistant chicks. a,b In the same row, values with different letters are significantly different between two groups (p < 0.05).
Table 2. Jejunum morphology of S. enteritidis-susceptible and -resistant chicks 1.
| Items |
Group S 2 |
Group R 2 |
p-Value |
| Villus height (μm) |
1084.62 ± 35.20 |
1125.93 ± 90.23 |
0.683 |
| Crypt depth (μm) |
149.56 ± 7.48 |
131.55 ± 16.28 |
0.348 |
| Ratio of villus height-to-crypt depth |
7.32 ± 0.34 |
8.94 ± 0.75 |
0.090 |
| Muscle thickness (μm) |
117.86 ± 7.09 |
118.02 ± 14.37 |
0.992 |
1 Results are expressed as means ± SEM, with n = 8 per group. 2 Group S = selected S. enteritidis-susceptible chicks; Group R = selected S. enteritidis-resistant chicks.
The intestinal mucosal barrier serves as the first line of defense between the host and the luminal environment. Composed of epithelial cells and tight junctions, this barrier can prevent the entry of harmful substances, such as pathogens and toxins, into host tissues, organs, and circulating blood
[27]. The intestinal epithelium is involved in the formation of the intestinal mucosal barrier by continuously secreting MUC2 to renew the intestinal mucosal layer. Impaired intestinal mucosal barrier function is a key determinant of the pathogenicity of some intestinal bacteria. Studies have shown that
Salmonella infection can disrupt the intestinal barrier of broilers, and promoting the expression of tight junction proteins through L-arginine supplementation can alleviate
Salmonella infection, indicating that there is a negative correlation between intestinal barrier function and the severity of
Salmonella infection
[28]. In this study, we compared the expression of genes encoding tight junction proteins and
MUC2 in
S. enteritidis-susceptible and
S. enteritidis-resistant chicks. The results showed that the mRNA expression of
occludin and
MUC2 in the jejunum of
S. enteritidis-resistant chicks was significantly higher than that of
S. enteritidis-susceptible chicks, further supporting that a negative correlation exists between intestinal mucosal barrier function and
S. enteritidis susceptibility.
Because proinflammatory cytokines are essential for initiating immune responses and eliminating pathogens in the host, we hypothesized that chicks in group R would exhibit higher levels of inflammation than those of group S, therefore explaining the greater resistance of the birds in group R to
S. enteritidis infection at the same dose of
S. enteritidis challenge. However, our results showed that there was no significant difference in the expression of most proinflammatory factor-related genes between the two groups. Furthermore, the gene expression of
iNOS and
IL6 showed the opposite trend to what would be expected, i.e., the expression of both genes was significantly higher in group S than in group R, whereas that of
IL10, coding for an anti-inflammatory factor, was significantly lower. These results suggested that inflammatory cytokines may play a role in the heterogeneous responses in an unexpected way. Or the higher expression levels of proinflammatory cytokine-related genes may also be considered to be a phenotype of
S. enteritidis-susceptible chicks, which is consistent with the results of the histopathological analysis of liver tissue. In addition, although
iNOS is believed to help cells resist bacterial invasion through the production of a large amount of NO, which serves as an antibacterial
[29], it is notable that the relationship between NO and
Salmonella in the host may not be merely antagonistic. It has been reported that
Salmonella needs NO as a nitrogen source for nitrate respiration, and a low NO concentration is indispensable for promoting
Salmonella growth
[30]. This may also explain why the invasion of
S. enteritidis in birds of group S was more severe, but their expression of the
iNOS gene were higher in our research.
In the chicken, the intestinal microbiota is composed of complex microbial communities that are involved in digestion and metabolism, the regulation of intestinal cells, vitamin synthesis, and the development and regulation of the host immune system
[31]. There is also accumulating evidence indicating that the intestinal microbiota profoundly influences the pathogenicity of
S. enteritidis [24]. Because the cecum is the most densely colonized microbial habitat in the chicken
[32], we systematically compared the cecal microbial composition of chicks from the different
S. enteritidis susceptibility groups. Alpha diversity refers to the richness and diversity within a microbial community in individual samples
[33], whereas beta diversity is a comparative analysis of microbial community composition in different samples. Although no significant difference was recorded for alpha diversity, significant differences in beta diversity were observed between the cecal samples of the two groups, which agreed with previous results showing that
Salmonella infection can lead to changes in cecal microbiota
[21].
The cecal microbial composition of the two groups at both the phylum and genus levels was analyzed using the Wilcoxon test. The results showed that at the phylum level, the relative abundance of
Acidobacteria,
Campilobacterota, and
Fusobacteriota were enriched in group S. The same results were obtained using LEfSe. At the genus level, 18 genera were identified as significantly differential microorganisms by the Wilcoxon test. Among them,
Fusobacterium,
Helicobacter, and
Butycicoccus were identified as marker microorganisms in group S using LEfSe. As we know,
Fusobacterium has been associated with gastric ulcers in pigs
[34] and colon carcinoma in humans
[35][36], and may represent a kind of new opportunistic pathogens of chickens worthy of further investigation
[37]. In addition, in the species level,
Helicobacter_pullorum has also been identified as a marker microorganism of group S, which is member of
Campilobacterota and a well-known zoonotic pathogen
[38]. These results revealed that chicks showing higher
S. enteritidis resistance has lower abundance of pathogenic bacteria in their cecal.
Furthermore, we identified a marker microorganism,
Desulfovibrio_piger, which was enriched in chicks of group R.
Desulfovibrio_piger, belonging to
Desulfovibrio spp., is a kind of sulfate reducing bacteria, which can functional reducing sulfate to hydrogen sulfide (H
2S) and plays an important role in intestinal hydrogen and sulfur metabolism. Although H
2S has been found to have dichotomous effects (stimulatory and inhibitory) on several gastrointestinal processes, it seems to be hazardous at high concentrations but favorable at low concentrations, and the overarching effect of H
2S appears to be beneficial. For example, H
2S can attenuate DSS-induced colitis, lessen the shortening of the colon lengths and colonic pathological damages, showing an overall protective effect in colitis via its anti-inflammatory properties
[39]. In addition, ATB-429, an H
2S releasing derivative of mesalamine, exhibits a marked increase in anti-inflammatory activity and potency in a murine model of colitis, as compared to mesalamine, seems promising in the treatment of inflammatory bowel disease
[40]. Our results were consistent with these above reports, as our chicks in group R showed higher abundance of
Desulfovibrio_piger and lower inflammation response at the same time. However, whether
Desulfovibrio_piger can really help chicks to resist the infection of
S. enteritidis by producing H
2S still need to be verified.
 +1 point
+1 point