1000/1000
Hot
Most Recent
 +1 point
+1 point
Immune checkpoint inhibitors (ICIs) improve the survival of patients with multiple types of cancer. However, low response rates and atypical responses limit their success in clinical applications. The paradoxical acceleration of tumor growth after treatment, defined as hyperprogressive disease (HPD), is the most difficult problem facing clinicians and patients alike. The mechanisms that underlie hyperprogression (HP) are still unclear and controversial, although a large number of studies have investigated the phenomenon and several associated factors have been reported. Gamma-interferon (IFN-γ) is a key cytokine in antitumor response and its levels increase during ICI therapy. Even though this factor has been widely associated with resistance to ICI therapy, it has not yet been demonstrated to be directly associated with HP. Nevertheless, data suggest that IFN-γ may contribute to HP onset through different mechanisms, including the activation of the inflammasome pathway, the expression of the immunosuppressive enzyme IDO1 and the triggering of activation-induced cell death (AICD) in effector T cells. These findings make IFN-γ worthy of attention in the context of HPD development.
Cancer immunotherapy aims to strengthen the immune system against tumors. The introduction of immunotherapy into clinical practice has provided clinicians with an innovative tool for the treatment of various solid and hematologic malignancies[1]. One of the most successful strategies is the administration of immune checkpoint inhibitors (ICIs), which are a wide range of monoclonal antibodies directed toward immune checkpoint (IC) proteins that are expressed on tumor cell and/or immune cell surfaces. The targeting of ICs reverses tumor-mediated immunosuppression and awakens immune responses[2]. The use of ICIs, either as monotherapy or combo-therapy, has shown favorable outcomes and remarkably long-term responses in patients with a large variety of cancer types, especially malignant melanoma and lung cancer[3][4][5]. The Food and Drug Administration (FDA) has so far approved seven ICIs that target cytotoxic T-lymphocyte antigen 4 (CTLA-4), programmed death-1 (PD-1) and PD-ligand 1 (PD-L1) for the treatment of several tumor types[5]. Nevertheless, response rates, according to the Response Evaluation Criteria in Solid Tumors (RECIST) version 1.1, for IC blockade in patients with solid tumors, range from 18 to 40% [6][7], as the beneficial clinical effects of ICI therapy are not long-lasting in some cases [8]. Moreover, the unique mechanism of action of ICIs can lead to unconventional responses, making IC blockade harmful to a subset of patients. Among these novel responses, the most relevant, in terms of negative clinical outcome, is hyperprogressive disease (HPD), a paradoxical acceleration of tumor growth induced by ICI therapy[8]. Even though several factors have been proposed as leading causes of HPD development, including alterations in T-cell subpopulations [9][10][11][12], tumor cells[13][14][15][16][17][18][19][20], cytokine secretion[21][22][23][24] and inflammation[25][26], the mechanisms underlying hyperprogression (HP) after ICI therapy remain unknown.
Gamma-interferon (IFN-γ) is a major regulatory and effector cytokine predominantly produced by T and natural killer (NK) cells in response to inflammatory and immune stimuli. In the tumor microenvironment (TME), IFN-γ, mainly produced by infiltrating T lymphocytes (TILs), is a key player in tumor immunosurveillance. The antitumor action includes antiproliferative, antiangiogenic and proapoptotic effects[27][28][29][30][31], in addition to the upregulation of major histocompatibility complex (MHC) class I molecules on tumor cells[32][33]. Moreover, IFN-γ activates CD8+ cytotoxic T lymphocytes, CD4+ Th1 cells, NK cells, dendritic cells (DCs) and macrophages, and stimulates the latter to switch towards the tumoricidal and proinflammatory M1 phenotype[33][34][35]. Conversely, IFN-γ also inhibits regulatory T (Treg) cell differentiation and function[36].
On the other hand, IFN-γ exerts a paradoxical immunosuppressive role that supports tumor progression and dissemination[37][38][39]. For instance, the activation of the IFN-γ receptor (IFNGR) on tumor cells activates the JAK/STAT signaling pathway, resulting in PD-L1 upregulation[40][41][42]. Nevertheless, alterations in the JAK/STAT pathway have been frequently associated with resistance to ICI therapy[43][44][45]]. Human melanoma cell lines with loss-of-function mutations in either JAK1 or JAK2 do not express IFN-γ-response genes after IFN-γ exposure. The analysis of the transcriptome of advanced melanoma under ICI therapy has highlighted an association between clinical response to treatment and the expression of IFN-γ-response genes involved in MHC class I and II upregulation[46]. Resistance to ICI therapy may therefore be due both to the incapacity of tumor cells to induce the full set of IFN-γ-response genes and to the loss of sensitivity to IFN-γ signaling. Interestingly, prolonged IFN-γ receptor signaling in tumor cells can also mediate resistance to ICIs through epigenomic changes in the JAK/STAT pathway[47]. IFN-γ can even support tumorigenesis by influencing the TME. The IFN-γ signaling pathway can indeed increase angiogenesis in the TME by inhibiting the expression of vascular endothelial growth inhibitor (VEGI) [48]. IFN-γ also suppresses the action of immune effector cells via the upregulation of immunosuppressive cytokines, including IL-21, IL-27 and IL-35[49][50][51][52][53][54], and by the recruitment and differentiation of Treg cells and myeloid-derived suppressor cells (MDSCs)[55][56][57][58][59][60][61][62][63].
All of the above-reported examples highlight the role that IFN-γ plays in tumor resistance to ICI therapy. Based on this evidence, it is reasonable to assume that IFN-γ is worthy of investigation in the context of HPD.
Several studies reported a negative correlation between MDSCs and the response to ICIs[64][65][66], leading to the suggestion that MDCSs can be a negative predictive marker for ICI therapy[67]. A few case reports noted a correlation between HPD development and the number of MDSCs in the peripheral blood and in the TME of patients[68][69]. Moreover, the recruitment of granulocytic MDSCs to the TME following ICI therapy, through the IFN-γ-dependent activation of the inflammasome pathway in cancer cells, has been reported[70]. PD-L1 upregulation by IFN-γ after ICIs and the consequent activation of the PD-L1 intrinsic signaling pathway in tumor cells trigger the activation of NLR family pyrin domain containing 3 (NLRP3), which leads to the downstream activation of the heat-shock proteins 70 (HSP70)/Toll-like receptor 4 (TLR4) signaling pathway and Wnt5a production. This signaling cascade ultimately leads to C-X-C motif chemokine ligand 5 (CXCL5) release, resulting in chemokine-dependent recruitment of polymorphonuclear-like MDSCs[70]. PD-L1 triggers NLRP3 activation by repressing STAT3, which is a transcription factor involved in IFN-cytotoxicity. Mutations in the intracytoplasmic DTSSK domain of PD-L1, which is a conserved sequence that acts as a negative regulator of PD-L1 functions, lead to hyperactive PD-L1 molecules in human tumors, enhancing the capacity of the PD-L1 intracellular pathway to interfere with STAT3 expression and phosphorylation[19]. Thus, in patients carrying mutations in the intracytoplasmic domain of PD-L1, the molecules of the inflammasome may be further augmented by PD-L1 upregulation after IFN-γ secretion in response to ICIs, eventually leading to HPD. Interestingly, a mutational analysis performed on tumors after pembrolizumab treatment highlighted the presence of missense or indel mutations in genes involved in the negative regulation of NLRP3 activation and inflammasome pathway[69], including the caspase recruitment domain (CARD8 and CARD11), protein flightless-1 homolog (FLII) and nuclear factor erythroid 2-related factor 2 (NFE2L2)[71][72][73][74]. Moreover, hyperprogressive tumors show mutations in NOTCH1, which seems to be involved in NLRP3 activation and inflammasome pathway [13][69][75][76][77]. In addition, the mechanism of MDSC recruitment in HPD may also be related to the impairment of effector T-cell activity, resulting in the expansion of Treg cells, the inhibition of NK cells and the secretion of immunosuppressive cytokines[78]. On the basis of the above-reported evidence, it can therefore be stated that IFN-γ-mediated recruitment of MDSCs in the TME may be a relevant aspect of HPD development.
Paracrine Wnt5a signaling is also involved in DC upregulation and enzymatic activity of indoleamine 2,3-dioxygenase (IDO1), promoting DC-mediated Treg differentiation[79][80]. IDO1 is a cytosolic enzyme that contributes to immune regulation by inducing metabolic changes in the local microenvironment. The enzyme catalyzes the rate-limiting step of tryptophan metabolism, which converts tryptophan (trp) into the downstream catabolite kynurenine (kyn). IDO1 is physiologically expressed by professional antigen-presenting cells (APCs), as well as by epithelial cells, the vascular endothelium and peripheral lymphoid organs, and acts as a peripheral IC, contributing to host defense against infection, peripheral immune tolerance, inhibition of local inflammation and autoimmunity [81][82]. Tumor cells use IDO1 expression as a mechanism of immune escape[83][84][85]. Trp depletion indeed results in a blockade of T-cell protein synthesis[86][87], while kyn and its derivatives induce PD-1 expression on activated T cells, together with the differentiation of Treg cells and tolerogenic DCs through aryl hydrocarbon receptor (AhR) activation[88][89][90][91]. In addition, IDO1 expression is induced, as is that of PD-L1, when some degree of inflammation occurs in the tumor, e.g., the presence of proinflammatory mediators, such as IFN-γ, as a mechanism of adaptive resistance against infiltrating T cells[92][93][94]. This aspect clearly has negative implications for ICI therapy, since increased IDO1 activity may increase tumor-infiltrating Treg cells, decrease TILs and accelerate tumor growth[95][96][97][98][99]. In non-small-cell lung cancer (NSCLC) patients treated with nivolumab, serum kyn/trp ratio was higher in early progressors with intrinsic resistance to anti-PD-1 therapy. Moreover, patients with high kyn/trp ratios showed a progression-free survival of three months, which is very similar to that of hyperprogressive patients in a study by Champiat et al. [100][101]. This evidence suggests that IDO1 induction by IFN-γ after ICI therapy may counteract the effectiveness of an otherwise beneficial treatment. Combination treatment with ICIs and IDO1 inhibitors in preclinical studies has been observed to enhance the infiltration and proliferation of effector T cells in the TME[102][103]. However, despite encouraging clinical results in early phase trials[104][105][106], in a randomized phase III study, patients with metastatic melanoma treated with both the IDO1 inhibitor epacadostat and pembrolizumab, had no benefit from the combined therapy, in comparison to pembrolizumab monotherapy[107]. This negative result may, however, be caused by limited preclinical data and either the incomplete inhibition of IDO1 or the compensatory expression of other trp-degrading enzymes[108]. Further clinical studies are therefore needed to understand whether this combined therapy may have therapeutic potential.
Interestingly, IDO1 upregulation is inversely correlated with p53[109], whose expression can be suppressed by IDO1 via the c-Jun N-terminal kinase (JNK) pathway[110]. It is notable that the downregulation of p53 has already been suggested as a HPD mechanism in patients with mouse double minute homolog 2 (MDM2) amplification[13]. Moreover, IDO1 expression is modulated by transforming growth factor-beta (TGF-β) via the Fyn-dependent phosphorylation of immunoreceptor tyrosine-based inhibition motifs (ITIMs) in IDO1, and the activation of the NF-κB pathway, which lead to a tolerogenic phenotype in DCs[111]. TGF-β can also activate the JNK pathway through TGF-β activated kinase 1 (TAK1) and c-Jun phosphorylation[112][113]. It is interesting to note that the TGF-β signaling pathway has been found to be transcriptionally upregulated in HPD tumors following IC blockade, as compared to treatment-naive tumors[69]. Therefore, HPD patients with IDO1-expressing tumors may present a hyperactivated JNK pathway which may result in p53 suppression. Moreover, post-therapy HPD tumors have also displayed transcriptional upregulation of PI3K/AKT and MAPK/ERK pathways, and it has been shown that PI3K and MAPK oncogenic mutations can favor constitutive IDO1 expression on tumor cells[69][114][115].
A further hypothesis that involves IFN-γ in HPD development, focuses on the differential immunological actions of IC blockade that occur depending on tumor burden. The combination of anti-CTLA-4 and anti-PD-1 therapy in mice with high tumor burden (HTB) leads to improved tumor control and to the generation of more activated antigen-specific T cells, as compared to mice with low tumor burden (LTB), in which combination treatment has been shown to compromise antitumor immune response, inducing the loss of antigen-specific T cells[116]. This finding was supported by retrospective clinical data from metastatic melanoma patients who received either monotherapy or combination therapy. Patients treated with dual IC blockade showed significantly lower response rates than those treated with monotherapy in low disease settings, but not in higher disease settings. The detrimental effect of combined therapy in the LTB state was associated with higher IFN-γ production, which was responsible for tumor-reactive CD8+ T-cell apoptosis via activation-induced cell death (AICD). AICD physiologically takes place in the early CD4+ and CD8+ T-cell priming stage, and leads to cell apoptosis to prevent immune hyperactivation. IFN-γ signaling is the key factor in activating this process, together with IL-2[117]. The induction of IFN-γ secretion after dual-blockade treatments can promote the apoptosis of tumor-reactive CD8+ T cells in the LTB setting, limiting the formation of effector memory antitumor responses. In the HTB state, prolonged antigen exposure may lead to T cells with a markedly exhausted phenotype, which may be more prone to reinvigoration after IC blockade. By contrast, in LTB or in the early tumor setting, short-term antigen exposure may be unable to induce a fully exhausted phenotype in T cells. Therefore, the activation of T-cell-receptor signaling against tumor antigens, in combination with dual IC blockade, may result in immune hyperactivation, triggering the AICD process. These findings appear to indicate that the paradoxical effect of IFN-γ in tumor response might derive from the differential exhaustion status of T cells in response to ICIs. Although the mechanisms underlying AICD have not yet been fully understood, Fas seems to be the major death receptor responsible for triggering the AICD pathway in CD4+ T cells[118]. Moreover, STAT1 and caspase 8, which are activated by the IFN-γ pathway, may be involved in the process[117]. The possible role of the activation of the AICD pathway by IFN-γ in HPD is supported by a study in which two hyperprogressive patients displayed depletion of the immune-cell populations involved in tumor clearance, including monocytes, central memory CD4+ T cells, NK cells and activated DCs[69]. In these patients, anti-PD-1 therapy had probably induced an accelerated AICD process in the antitumor activating lymphocytes, as suggested by the activation of apoptosis gene sets and the upregulation of marker genes in the bcl-2 pathway after treatment. These studies suggest that, in some patients, ICI therapy may be responsible for excessive activation of the immune response, which could trigger regulatory mechanisms and hinder therapeutic antitumor effects.
In conclusion, it may be suggested that IFN-γ contributes to HPD onset in predisposed patients via the induction of the inflammasome pathway and consequent MDSC recruitment, the induction of IDO1 activity, which may result in the downregulation of p53 in tumor cells, and, finally, the activation of AICD, which leads to T-cell depletion (Figure 1).
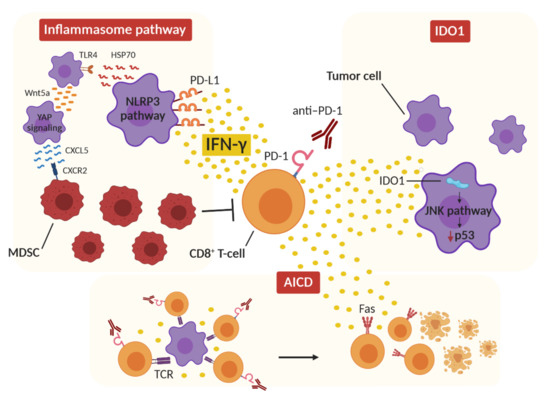
Figure 1. Proposed IFN-γ-dependent mechanisms of hyperprogression. The release of IFN-γ from CD8+ T cells after ICI therapy can activate the inflammasome pathway by upregulating PD-L1 expression on tumor cells and activating NLRP3 signaling, resulting in immunosuppressive MDSC recruitment in the tumor microenvironment. At the same time, IFN-γ can induce IDO1 activity in tumor cells, which activates the JNK pathway, leading to p53 downregulation and tumor growth. Finally, the concomitant stimulation of tumor-specific CD8+ T cells by ICI therapy and T-cell-receptor (TCR) activation results in a hyperactivated immune environment in which IFN-γ triggers the activation-induced cell death (AICD) mechanism and T-cell Fas-mediated apoptosis.





