1000/1000
Hot
Most Recent
 +1 point
+1 point
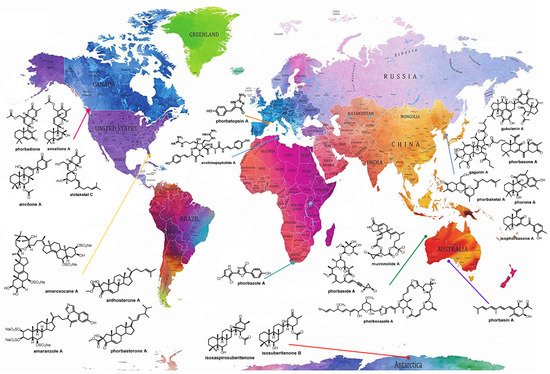
Porifera, commonly referred to as marine sponges, are acknowledged as major producers of marine natural products (MNPs). Sponges of the genus Phorbas have attracted much attention over the years. They are widespread in all continents, and several structurally unique bioactive compounds have been identified from this species.
| Name | Class | Species | Cell Lines | Dose/Concentration | Reference | |
|---|---|---|---|---|---|---|
| 13 | Zarzissine | Alkaloid | Phorbas tenacior |
P-388 a | IC50 12 µg/mL | [22] |
| KB b | IC50 5 µg/mL | |||||
| NSCLC-N6 c | IC50 10 µg/mL | |||||
| 17 | Phorboxazole A | Macrolide | Phorbas sp. | HCT-116 d | GI50 4.36 × 10−10 M | [23] |
| HT29 d | GI50 3.31 × 10−10 M | |||||
| 19 | Muironolide A | Macrolide | Phorbas sp. | HCT-116 d | IC50 96.5 μg/mL | [24] |
| 20 | Phorbaside A | Macrolide | Phorbas sp. | HCT-116 d | IC50 30.0 μM | [25] |
| 22 | Phorbaside C | Macrolide | Phorbas sp. | HCT-116 d | IC50 2 μM | [25] |
| 23 | Phorbaside D | Macrolide | Phorbas sp. | HCT-116 d | IC50 61.9 μM | [25] |
| 24 | Phorbaside E | Macrolide | Phorbas sp. | HCT-116 d | IC50 10.2 μM | [25] |
| 29 | Phorbasterone A | Steroid | Phorbas amaranthus |
HCT-116 d | IC50 1–3 µg/mL | [26][27] |
| 30 | Phorbasterone B | Steroid | Phorbas amaranthus |
HCT-116 d | IC50 1–3 µg/mL | [26][27] |
| 31–32 | Phorbasterone C | Steroid | Phorbas amaranthus |
HCT-116 d | IC50 1–3 µg/mL | [26][27] |
| 33–34 | Phorbasterone D | Steroid | Phorbas amaranthus |
HCT-116 d | IC50 1–3 µg/mL | [26][27] |
| 45 | Phorbaketal A | Sesterterpenoid | Phorbas sp. | A549 c | IC50 11–12 µg mL−1 | [28][29] |
| HT-29 d | IC50 11–12 µg mL−1 | |||||
| HepG2 e | IC50 11–12 µg mL−1 | |||||
| 46 | Phorbaketal B | Sesterterpenoid | Phorbas sp. | A549 c | IC50 12–460 µg/mL | [28][29] |
| HT-29 d | IC50 12–460 µg/mL | |||||
| HepG2 e | IC50 12–460 µg/mL | |||||
| 47 | Phorbaketal C | Sesterterpenoid | Phorbas sp. | A549 c | IC50 12–460 µg/mL | [28][29] |
| HT-29 d | IC50 12–460 µg/mL | |||||
| HepG2 e | IC50 12–460 µg/mL | |||||
| HT-29 d | LG50 5–15 μM | |||||
| 50 | Phorbaketal N | Sesterterpenoid | Phorbas sp. | PANC-1 f | IC50 11.4 µM | [30] |
| A498 g | IC50 18.7 µM | |||||
| ACHN g | LC50 24.4 µM | |||||
| 84 | Isosuberitenone B | Sesterterpenoid | Phorbas areolatus |
A549 c | IC50 8.8 μM | [31] |
| HT-29 d | IC50 9.0 μM | |||||
| HepG2 e | IC50 7.4 μM | |||||
| MCF-7 h | IC50 8.8 μM | |||||
| 85 | 19-episuberitenone B | Sesterterpenoid | Phorbas areolatus |
A549 c | IC50 5.1 μM | [31] |
| HT-29 d | IC50 6.4 μM | |||||
| HepG2 e | IC50 5.0 μM | |||||
| MCF-7 h | IC50 5.1 μM | |||||
| 88 | Phorbasin B | Diterpene | Phorbas sp. | A549 c | LG50 5–15 μM | [32] |
| HT-29 d | LG50 5–15 μM | |||||
| 89 | Phorbasin C | Diterpene | Phorbas sp. | A549 c | LG50 5–15 μM | [32] |
| HT-29 d | LG50 5–15 μM | |||||
| 91 | Phorbasin E | Terpenyl-taurine | Phorbas sp. | A549 c | LG50 5–15 μM | [32] |
| HT-29 d | LG50 5–15 μM | |||||
| 101 | Gagunin A | Diterpenoid | Phorbas sp. | K-562 a | LC50 50.1 µg/mL | [33] |
| 102 | Gagunin B | Diterpenoid | Phorbas sp. | K-562 a | LC50 10.4 µg/mL | [33] |
| 103 | Gagunin C | Diterpenoid | Phorbas sp. | K-562 a | LC50 0.71 µg/mL | [33] |
| 104 | Gagunin D | Diterpenoid | Phorbas sp. | K-562 a | LC50 0.13 µg/mL | [33] |
| 105 | Gagunin E | Diterpenoid | Phorbas sp. | K-562 a | LC50 0.03 µg/mL | [33] |
| 106 | Gagunin F | Diterpenoid | Phorbas sp. | K-562 a | LC50 0.11 µg/mL | [33] |
| 107 | Gagunin G | Diterpenoid | Phorbas sp. | K-562 a | LC50 2.0 µg/mL | [33] |
| 108 | Gagunin H | Diterpenoid | Phorbas sp. | K-562 a | LC50 10.0 µg/mL | [34] |
| 109 | Gagunin I | Diterpenoid | Phorbas sp. | K-562 a | LC50 11.5 µg/mL | [34] |
| 110 | Gagunin J | Diterpenoid | Phorbas sp. | K-562 a | LC50 9.1 µg/mL | [34] |
| 111 | Gagunin K | Diterpenoid | Phorbas sp. | K-562 a | LC50 17.5 µg/mL | [34] |
| 112 | Gagunin L | Diterpenoid | Phorbas sp. | K-562 a | LC50 12.5 µg/mL | [34] |
| 113 | Gagunin M | Diterpenoid | Phorbas sp. | K-562 a | LC50 0.71 µg/mL | [34] |
| 114 | Gagunin N | Diterpenoid | Phorbas sp. | K-562 a | LC50 > 50 µg/mL | [34] |
| 115 | Gagunin O | Diterpenoid | Phorbas sp. | K-562 a | LC50 11.1 µg/mL | [34] |
| 116 | Gagunin P | Diterpenoid | Phorbas sp. | K-562 a | LC50 8.5 µg/mL | [34] |
| 117 | Gagunin Q | Diterpenoid | Phorbas sp. | K-562 a | LC50 > 50 µg/mL | [34] |
| 118 | Gukulenin A | tetraterpenoid | Phorbas gukulensis |
HCT-116 d | IC50 62 nM | [35] |
| FaDu b | IC50 57 nM | |||||
| SN12C g | IC50 92 nM | |||||
| MKN45 j | IC50 0.13 nM | |||||
| TOVG-21G i | IC50 0.04 μM | [36] | ||||
| OVCAR-3 i | IC50 0.13 μM | |||||
| A2780 i | IC50 0.03 μM | |||||
| SKOV3 i | IC50 0.36 μM | |||||
| 119 | Gukulenin B | tetraterpenoid | Phorbas gukulensis |
HCT-116 d | IC50 0.55 μM | [35] |
| A2780 i | ||||||
| FaDu b | IC50 0.63 μM | |||||
| SN12C g | IC50 0.61 μM | |||||
| MKN45 j | IC50 0.72 μM | |||||
| 123 | Gukulenin F | Tetraterpenoid | Phorbas gukulensis |
K-562 a | LC50 0.4 µM | [35] |
| FaDu b | IC50 0.63 μM | |||||
| SN12C g | IC50 0.61 μM | |||||
| MKN45 j | IC50 0.72 μM |
| Compound | Class | Species | Biological Activity | Dose/Concentration | Reference | |
|---|---|---|---|---|---|---|
| 1 | Anchinopeptolide A | Alkaloid | P. tenacior | Displacement of specific ligands from their biochemical receptors | 5 µg/mL-average inhibition values roughly 35–40% in all receptor binding | [37][38] |
| 2 | Anchinopeptolide B | Alkaloid | P. tenacior | Displacement of specific ligands from their biochemical receptors | 5 µg/mL-71% human B2 bradykinin receptor; 80% neuropeptide Y receptor | [37][38] |
| 3 | Anchinopeptolide C | Alkaloid | P. tenacior | Displacement of specific ligands from their biochemical receptors | 5 µg/mL-62% somatostatin receptor; 52% human B2 bradykinin receptor; 57% neuropeptide Y receptor | [37][38] |
| 4 | Anchinopeptolide D | Alkaloid | P. tenacior | Displacement of specific ligands from their biochemical receptors | 5 µg/mL-77% somatostatin receptor | [37][38] |
| 13 | Zarzissine | Alkaloid | P. topsenti | Antimicrobial | Paper disk agar-(100 µg, purified product) 12,10, and 11 mm | [22] |
| 14 | p-Hydroxybenzaldehyde | Alkaloid | P. topsenti | Antimicrobial | Paper disk agar-(100 µg, purified product) 8,7, and 7 mm | [22] |
| 14 | Phorbatopsin A | Alkaloid | P. topsenti | Antioxidant | ORACFL 0.88 ± 0.28 | [39] |
| 15 | Phorbatopsin B | Alkaloid | P. topsenti | Antioxidant | ORACFL 0.50 ± 0.08 | [39] |
| 16 | Phorbatopsin C | Alkaloid | P. topsenti | Antioxidant | ORACFL 0.21 ± 0.02 | [39] |
| 17 | Phorboxazole A | Macrolide | Phorbas sp. | Antifungal | Agar disk diffusion assay-C. albicans: 12 mm (1 µg) and 9 mm (0.1 µg); Saccharomyces carlsbergensis: 1, 20 mm (1 µg), and 13 mm (0.1 µg) | [23] |
| 18 | Phorboxazole B | Macrolide | Phorbas sp. | Antifungal | Agar disk diffusion assay-C. albicans: 11 mm (1 µg) and 8 mm (0.1 µ g); Saccharomyces carlsbergensis: 1, 16 mm (1 µg), and 10 mm (0.1 µg) | [23] |
| 19 | Muironolide A | Macrolide | Phorbas sp. | Antifungal | MIC 16 μg/mL | [24] |
| 22 | Phorbaside C | Macrolide | Phorbas sp. | Macrophage infectivity potentiator (Mip) | Binding affinity of 75 with Chlamydia pneumoniae | [40] |
| 37 | Amaroxocane A | Steroid | P.amaranthus | Anti-predatory activity | Little feeding deterrence (8/10 pellets eaten) | [41] |
| 38 | Amaroxocane B | Steroid | P. amaranthus. | Anti-predatory activity | Significant deterrent activity (3/10 pellets eaten) | [41] |
| 45 | Phorbaketal A | Sesterterpenoid | Phorbas sp. | Osteogenic differentiation Anti-inflammatory |
Phorbaketal A stimulates TAZ-mediated osteoblast differentiation through the activation of extracellular signal-regulated kinase (1–10 µg/mL) Dose dependent inhibition of LPS-induced production of inflammatory cytokines and the transcriptional activity NF-κB (2.5, 5, and 10 μM) and adipocyte differentiation through transcriptional coactivator with PDZ-binding motif (1–10 µg/mL) |
[42] [43][44] |
| 60 | Alotaketal A | Sesterterpenoid | Phorbas sp. | cAMP signaling activation | cAMP cell signaling pathway-EC50 of 18 nM |
[45] |
| 61 | Alotaketal B | Sesterterpenoid | Phorbas sp. | cAMP signaling activation | cAMP cell signaling pathway-EC50 of 240 nM |
[45] |
| 62 | Alotaketal C | Sesterterpenoid | Phorbas sp. | Latency-reversing agent (LRA) | HIV-1 provirus/GFP expression of J-Lat 9.2 cells-1 μM | [46][47][48] |
| 63 | Alotaketal D | Sesterterpenoid | Phorbas sp. | Latency-reversing agent (LRA) | HIV-1 provirus/GFP expression of J-Lat 9.2 cells-30 μM | [46][47][48] |
| 65 | Ansellone A | Sesterterpenoid | Phorbas sp. | cAMP signaling activation Latency-reversing agent (LRA)cAMP activator |
cAMP cell signaling pathway-EC50 of 14 µM HIV-1 provirus/GFP expression of J-Lat 9.2 cells-30 μM |
[47][49] |
| 66 | Ansellone B | Sesterterpenoid | Phorbas sp. | Inhibition of inducible NOS (iNOS) | RAW 264.7 LPS-activated mouse macrophage cells-IC50 = f 4.5 μM, | [50] |
| 73 | Anvilone A | Sesterterpenoid | Phorbas sp. | Latency-reversing agent (LRAs) | HIV-1 provirus/GFP expression of J-Lat 9.2 cells-30 μM | [47] |
| 76 | Phorbasone A | Sesterterpenoid | Phorbas sp. | Osteogenic properties | Calcium deposition effect at a concentration of 0.5 μg/mL | [51] |
| 79 | Phorbasone A acetate | Sesterterpenoid | Phorbas sp. | Inhibition of inducible NOS (iNOS) | Inhibitory activity on NOS in RAW 264.7 LPS-activated mouse macrophage cells-IC50 = 2.8 μM | [50] |
| 83 | Oxaspirosuberitenone | Sesterterpenoids | P. areolatus | Antimicrobial | Activity against MRSA at the highest concentration tested (160 µM) | [31] |
| 94 | Phorbasin H | Diterpenoid | Phorbas sp. | Antifungal | Suppression of the hyphal development of C. albicans (250 μg/mL) | [52] |
| 93–95 | Phorbasins | Diterpenoid | Phorbas sp. | Antifungal | EtOH extract-growth inhibitory activity against the gram-positive bacteria Staphylococcus aureus and Micrococcus luteus–Concentration: N/A | [53][54] |
| 104 | Gagunin D | Diterpenoid | Phorbas sp. | Anti-melanogenic | IC50 = 5.7 µg/mL; 10 µM on UVB irradiated human skin models demonstrated a considerable reduction melanin biosynthesis | [55] |
| 101–117 | Gagunins | Diterpenoid | Phorbas sp. | Isocitrate lyase (ICL) inhibition | LC50 of 55–140 µg/mL | [34] |
| 125 | Astaxanthin | Carotenoid | P. topsenti | Antioxidant | ORACFL 0.22 ± 0.02 | [39] |
| 126 | Adonirubin | Carotenoid | P. topsenti | Antioxidant | ORACFL 0.024 ± 0.001 | [39] |
| 127 | Taurine | Sulfonic acid | P. topsenti | Antioxidant | ORACFL 0.083 ± 0.013 | [39] |
| 128 | Taurobetaine | Sulfonic acid | P. topsenti | Antioxidant | ORACFL 00.019 ± 0.002 | [39] |





