Autophagy is a stress-responsive catabolic process that degrades intracellular components through lysosomal enzymes
[36]. In normal physiological state, only a small amount of autophagy occurs in cells to maintain homeostasis. When cells are stimulated by intracellular and extracellular factors such as starvation, hypoxia, pathogen invasion, etc.
[37], a large number of autophagy can be induced through the transduction of cell signaling pathways
[38]. Thus, autophagy is a pivotal actor in development, immune response, as well as metabolic regulation and has been shown to be associated with cellular modifications related to senescence, with most studies now suggesting that a reduced autophagic potential is one of the factors of cell senescence
[36][39][40]. Autophagy not only removes protein aggregates, but also damages organelles and plays a role in quality control of the cytoplasm such as mitophagy
[41]. In the case of damaged mitochondria, mitophagy removes malfunctioning mitochondria to maintain the population at an optimal state. In recent years, mitophagy has received increasing attention since mitochondrial dysfunction is at the foundation of numerous diseases and a growing number of studies also suggested mitophagy as a therapeutic target
[42][43].
Autophagy is composed of several closely related steps including autophagy initiation, autophagosome maturation, and autophagolysosome fusion, which involves many important autophagy-related proteins and complexes
[44]. These core autophagy proteins include the following parts: (1) the ATG1/ULK1 complex, including ATG1, ATG11, ATG13, ATG17, ATG29, and ATG31, is the only core protein complex with serine/threonine kinase activity in autophagy signaling pathway. The ULK1 complex acts as a bridge connecting upstream energy receptors mTOR and AMPK with downstream autophagosomes in vivo
[45][46], and plays an important role in autophagy initiation
[47][48]; (2) the PI3K complex, including Vps34, Vps15, ATG6/Beclin1, and ATG14, catalyze the conversion of the lipid molecule PI to PI3P, thereby recruiting the protein that binds to PI3P
[49]. Vps34 is the class III PI3K in mammals. Vps34 is activated by binding to Vps15 and further binds to Beclin1 to form the Vps34-Vps15-Beclin1 complex. During autophagy, Vps34-Vps15-Beclin1 binds to a variety of autophagy-related proteins. For example, ATG14 is combined with to Vps34-Vps15-Beclin1 to form ATG14-Vps34-Vps15-Beclin1, which is involved in the formation of autophagic vesicles
[50]; the (3) ATG9 and WIPI/ATG2-ATG18 complex. ATG9 is a transmembrane protein with six transmembrane domains, which may play a role in regulating autophagy by affecting vesicular transport
[51]. ATG9 circulates in autophagic vesicles and cytoplasm, depending on ATG17 or ATG11 complex to locate PAS, and ATG2-ATG18 complex to leave PAS
[52][53]. In mammals, specific silence of mATG9 gene can inhibit the formation of autophagic vesicles and protein degradation, and inhibit the occurrence of autophagy; and (4) ubiquitin-like systems ATG12-ATG5 and ATG8/LC3. There are two ubiquitin-like binding pathways involved in autophagosome formation. Both ATG8 and ATG12 are ubiquitin-like proteins, ATG12 can covalently bind with ATG5, and ATG8 can covalently bind with the lipid molecule phosphatidylethanolamine PE
[54][55]. Similar to the ubiquitin system, ATG12 is transmitted by ATG7 to ATG10, which eventually binds to the lysine side chain of ATG5 and forms a complex with ATG16, promoting the exposure of membrane-binding sites on ATG5. Similarly, ATG7 transfers ATG8 to ATG3
[56]. With the help of ATG12-ATG5-ATG16, LC3 conjugates to lipid molecule phosphatidylethanolamine (PE), promoting isolation membrane expansion and autophagic vesicle completion
[57]. These key proteins in the complex autophagy regulatory network are regulated by a variety of molecular signals, including ubiquitin ligases, deubiquitinases and miRNAs
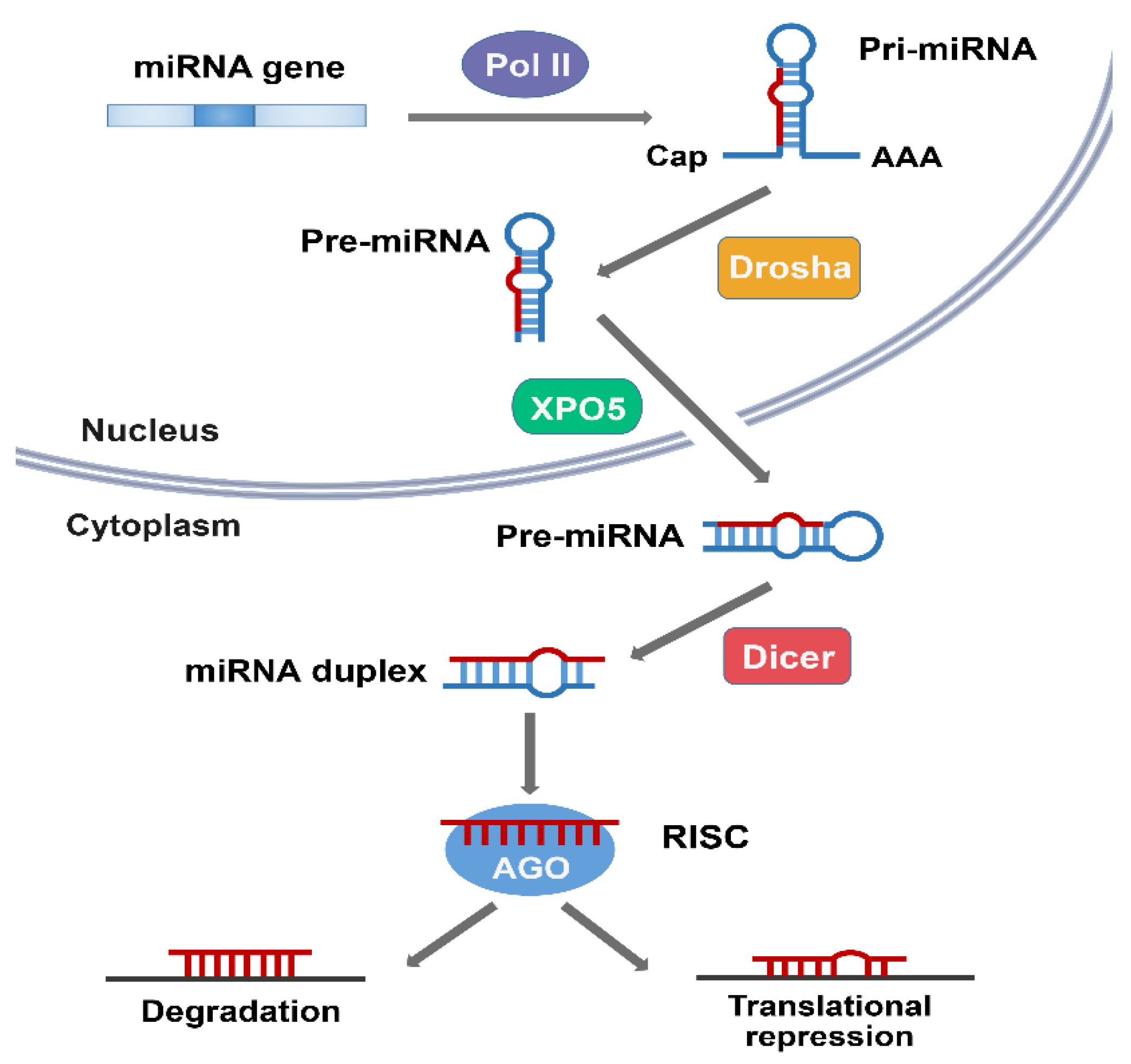
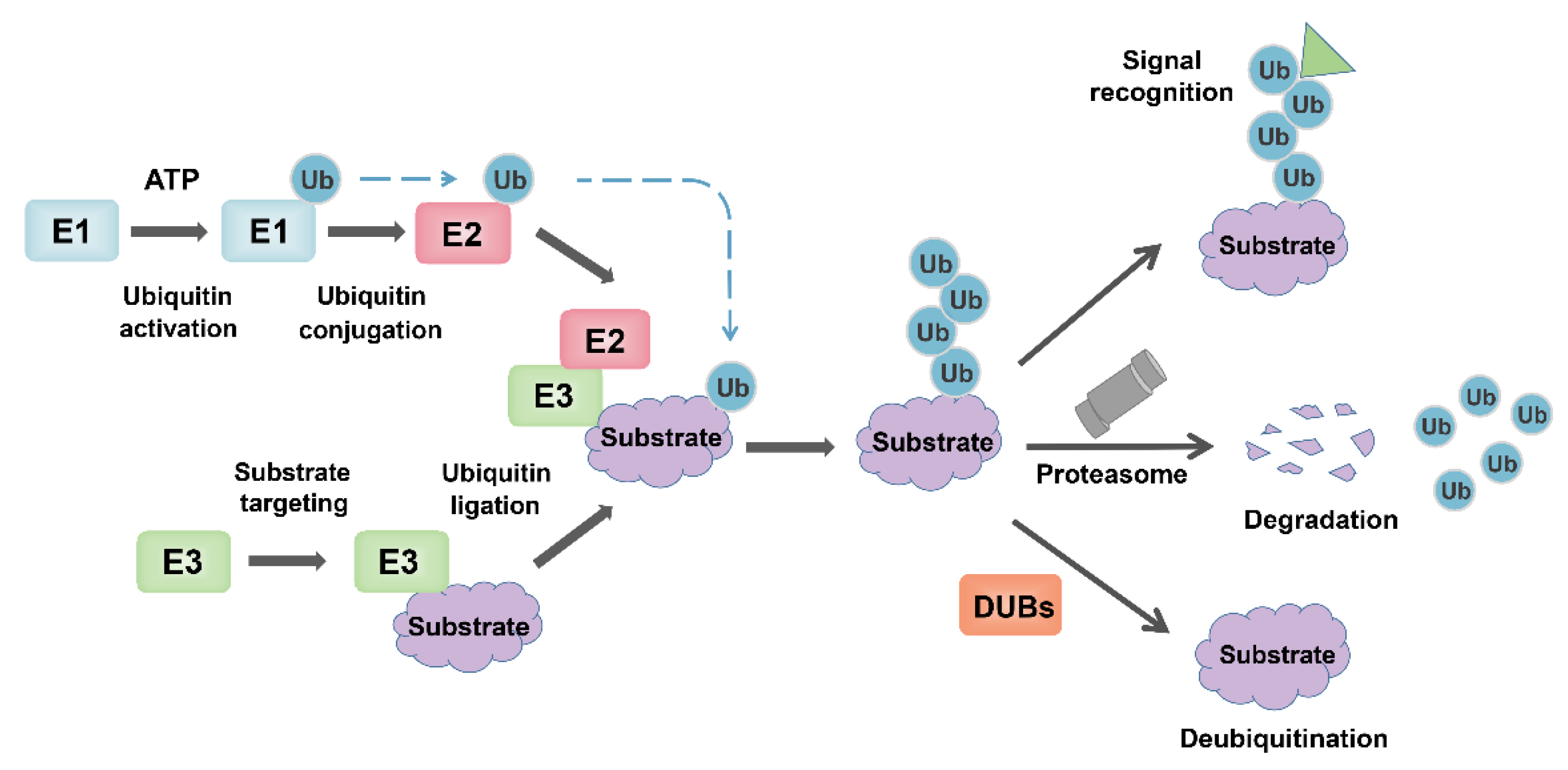
[58].
 +1 point
+1 point