The aging of bone marrow (BM) remains a very imperative and alluring subject, with an ever-increasing interest among fellow scientists. A considerable amount of progress has been made in this field with the established ‘hallmarks of aging’ and continued efforts to investigate the age-related changes observed within the BM. Inflammaging is considered as a low-grade state of inflammation associated with aging, and whilst the possible mechanisms by which aging occurs are now largely understood, the processes leading to the underlying changes within aged BM remain elusive. The ability to identify these changes and detect such alterations at the genetic level are key to broadening the knowledgebase of aging BM. Next-generation sequencing (NGS) is an important molecular-level application presenting the ability to not only determine genomic base changes but provide transcriptional profiling (RNA-seq), as well as a high-throughput analysis of DNA–protein interactions (ChIP-seq). Utilising NGS to explore the genetic alterations occurring over the aging process within alterative cell types facilitates the comprehension of the molecular and cellular changes influencing the dynamics of aging BM.
1. Introduction
Aging is a complex process impacting the lives of many people in their 60s and above by means of reduced mobility, increased vulnerability to diseases and poor quality of life (QOL). Biologically, this results from a wide range of cellular and molecular changes over a prolonged period of time
[1]. The global population of individuals over the age of 65 was 524 million in 2010, and the number is projected to be 1.5 billion by 2050
[2]. Considering that advancing age is often associated with several diseases
[3], the World Health Organization (WHO) predicts that the economic and social burdens associated with these diseases are rising sharply with older age
[2] and, thus, are often referred to as age-related diseases (ARDs).
As a field with immense growing interest, the last few decades have seen a rise in exploring the theories and mechanisms that lead to age-related changes and ARDs. Lopez-Otin et al. set the benchmark with their ‘
The hallmarks of aging’ article discussing the underlying factors and mechanisms that lead to changes in the elderly
[4]. Damage to the DNA or the DNA damage responses (DDR) are among the oldest mechanisms accepted for age-related changes. This refers to the accumulation of damage to the DNA or mitochondrial DNA (mtDNA) due to various factors over time that ultimately leads to alterations in the chromosomes and overall genetic instability
[5]. Another mechanism observed frequently in aging tissues is the presence of reactive oxygen species (ROS), also known as the free radical theory
[6], that have an unpaired electron in their outermost shell, making them unstable. In both healthy and young individuals, antioxidants inherently present within the body balance the presence of ROS. However, with advancing age, the balance is reduced, causing the oxidation of nucleic acids and DNA damage
[4]. The shortening of the telomere length (or telomere attrition) with every cell division is an additionally observed mechanism contributing to age-related changes
[7]. Furthermore, stem cell exhaustion or the changes deciphered in the number and functions of stem cells also contribute to age-related changes and ARDs. Stem cell exhaustion has also been linked to the other mechanisms of aging and is potentially caused by the previous mechanisms mentioned above
[8]. Whilst proteostasis (or protein homeostasis) serves as the quality control for maintaining the proteome, this function is known to decline with age
[8]. The loss of proteostasis leads to the presence and accumulation of misfolded proteins that have been linked to ARDS
[4][9], making it another hallmark of aging. Finally, changes in the DNA methylation patterns, post-translational histone modifications and chromatin remodelling are epigenetic alterations that can additionally be held accountable as mechanisms contributing to age-relate changes
[10].
More recently, aging has been identified as a state of ‘chronic, low-grade, sterile inflammation’ with poorly understood mechanisms. This state is known as ‘inflammaging’ and has been associated with multiple ARDs that contribute to morbidity and mortality in the elderly
[11]. Theoretically, inflammaging is an extension of the ‘theories of aging’ and essentially originates from the hypothesis that the complex process of aging and deterioration of health in ARDs must have a common origin
[11]. Considering that most ARDs, if not all of them, share an inflammatory pathogenesis, inflammaging is a significant risk factor to the health of the elderly
[12].
The lack of ability to fight off infections, cognitive impairment, frailty and immunosenescence are some of the most debilitating impacts of advancing age, causing significant challenges to the QOL within the elderly. Amongst these challenges faced by the elderly is reduced mobility due to alterations in the musculoskeletal system, including symptoms of pain in the joints, inflammation and increased vulnerability towards disorders like osteoarthritis and osteoporosis
[13]. Loss of muscle or sarcopenia, decreasing muscle strength, increased stiffness, pain in joints, thinning of cartilage and loss of bone density are widely recognised signs associated with age-related changes and ARDs in the musculoskeletal system
[14].
The BM is a critical part of the musculoskeletal system that is known for its dynamic nature in the human body. It is the origin of blood formation or haematopoiesis, serves as the reserve for the different stem cells, bone-forming cells, their progenitors, macrophages and adipocytes, alongside various growth factors, cytokines and other soluble factors
[15]. Changes with advancing age in the BM include altered cellularity, increased adipogenesis, myeloid skewing and decline in proliferation and regeneration potential
[15]. These changes have frequently been associated with diseases of higher risk like cancer, osteoarthritis and osteoporosis
[13]. Evidence suggests that there are similarities between haematopoietic alterations during inflammatory conditions (like infections) and during aging. These include dominant myelopoiesis in relation to immune senescence and impaired B-lymphopoiesis. This is accompanied by increased adipogenesis in the BM and the release of proinflammatory cytokines, including IL-6, TNF-α and IL-1α, and C-reactive proteins. All of these observations provide evidence for the connections linking age-related changes and inflammaging
[16].
Considering that BM aging and the resulting changes may have life-altering effects, numerous techniques have been utilised to understand these age-related changes at the cellular level. The investigating parameters, including senescence, proliferation, telomere length, gene expression and reactive oxygen species (ROS), have all aided in the understanding we have today regarding BM aging, including the probable causes behind these mechanisms
[4][17]. However, a technique that can provide us with a large amount of data at the genetic level for the identification of new pathways would be ideal for evaluating age-related changes in the BM. Next-generation sequencing (NGS) provides us with exactly that, making it among the most desirable techniques for studying changes with advancing age
[18][19].
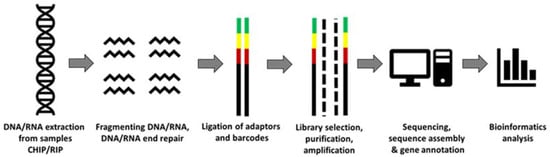
NGS has emerged from the initial sequencing of genomes performed using traditional Sanger sequencing and is thus referred to as second-generation sequencing technology
[18]. Fundamentally, the process involves randomly fragmenting DNA/RNA into smaller sizes (typically around 100–300 bp, depending on sequencing strategy), adaptor ligation and immobilisation of the given fragments to construct the DNA or cDNA libraries
[20][21]. The latter are lastly amplified to generate multiple copies of each fragment and then subjected to high-coverage sequencing using fluorescence or chemiluminescent-based methods (i.e., bridge amplification)
[22]. The resulting reads are finally mapped to the given reference genome of the species, and using assorted bioinformatics pipelines and computational processes, the DNA–RNA, RNA–protein and DNA–protein networks can be evaluated, providing an unparalleled ability to explore various molecular networks
[18]. This is briefly outlined in
Figure 1. Whilst Illumina, short read and sequencing-by-synthesis remains the ‘current gold standards of clinical and research sequencing’, the scope of NGS has massively broadened within the past five years
[23]. The emergence of Pacific Biosciences (PacBio) and Oxford Nanopore Technologies (ONT) long-read sequencing technology (permitting 15 kb-30-kb uninterrupted reads) is a significant contrast from the shorter read lengths provided by Illumina (i.e., 2 × 250 bp on Novaseq 6000™)
[24]. Thus, in the context of BM aging, both types of these highly accurate NGS platforms enhance our comprehension of the mutational and evolutionary developments through the understanding of further intricate forms of genetic variations
[23].
Figure 1. Basic next-generation sequencing (NGS) workflow adapted from Reference
[25].
2. Aging in the Bone Marrow
The BM serves as the integration between the skeleton and the marrow, providing a niche microenvironment for the cells and growth factors residing here. The BM is home to haematopoietic stem cells (HSCs), mesenchymal stem cells (MSCs) and lymphoid and myeloid progenitors, as well as endothelial cells
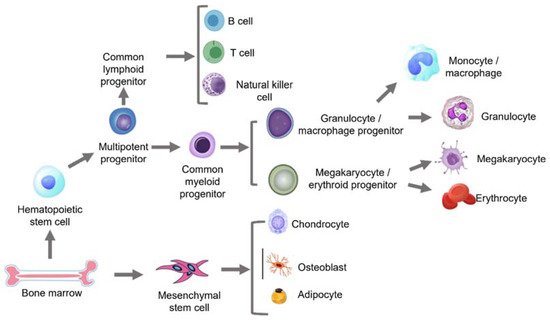
[26]. Stem cells give rise to different lineages (
Figure 2) and are known for their ability to regenerate. However, this ability declines with advancing age, which is now known as stem cell exhaustion, identified as one of the key hallmarks of aging
[4][17]. Whilst the different stem cells and immune cells all coexist within the BM niche, how their functions are altered with advancing age have been investigated separately
[19]. This section summarises the changes associated with aging amongst the various cell types and the BM in general.
Figure 2. An overview of the lineages arising from the HSC and MSC populations.
The bone marrow gives rise to both self-renewing HSCs and MSCs. HSCs further turn into multipotent progenitors that form common lymphoid progenitors (CLPs) and common myeloid progenitors (CMPs). CLPs finally form B cells, T cells and natural killer (NK) cells, while CMPs give rise to myeloid cell types that include the different types of granulocytes/macrophage progenitors and megakaryocyte/erythroid progenitors. MSCs differentiate into chondrocytes, osteoblasts and adipocytes, which eventually form the cartilage, bone and fat in the human body, as summarised in
Figure 2, which has been recreated and adapted from Wang et al.
[27].
2.1. The Aging Bone Marrow (BM) Niche
The BM niche is the unique microenvironment that is provided by the distinctive architecture of the bone, the cellular components and the growth factors and proteins within it. The two major types of stem cells, i.e., haematopoietic stem cells (HSCs) and mesenchymal stem cells (MSCs), give rise to different lineages and, thus, form an essential part of this niche
[28]. Being heterogenous in nature, the BM undergoes several alterations with advancing age, including changes in cellularity of the stem cells
[29] and loss of the ‘bone–fat balance’ within the BM
[30]. Age-related bone loss and decline in bone strength and density, followed by an increased vulnerability to BM and/or bone-related diseases like osteoarthritis, myelodysplastic syndrome and haematological malignancies, are also well-documented
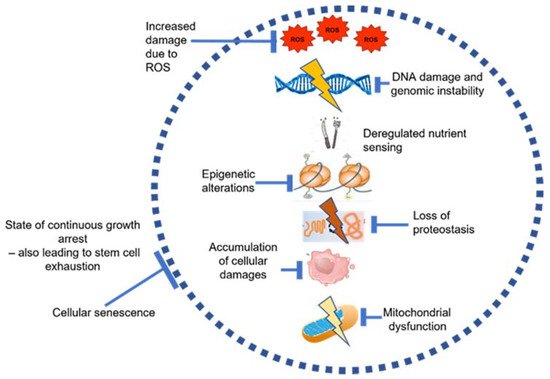
[16]. The underlying causative factors for these changes have been proposed to be a combination of several of the ‘hallmarks of aging’, including oxidative stress due to ROS, DNA damage, mitochondrial dysfunction and cellular senescence (
Figure 3)
[4][17].
Figure 3. Underlying causes of aging inside the BM, recreated and adapted using information from Lopez Otin et al.
[4] and González-Gualda et al.
[31].
More recently, Guidi et al. established that the aged BM niche ‘restrains rejuvenated HSC’. They found that using rejuvenated old HSCs upon transplanting into the BM of young mice was able to partly sustain youthful functions and suggested that age-related changes in stem cells potentially result from both cell intrinsic factors and extrinsic factors (niche) of the BM
[32]. Metabolically, an accumulation of free fatty acids (FFA) combined with the decline in the levels of nucleic acids and the amino acid pool has been documented, potentially due to the signature oxidative stress revealed in their transcriptome data associated with mitochondrial dysfunction
[33]. Apart from these, the process of myeloid skewing, a process where the lymphoid cells become more myeloid-oriented in the elderly, is also a key indicator of BM aging
[34].
Whether it is the altered microenvironment (cell extrinsic factors) leading to the reduced functionality of the cells within the BM with advancing age or the intrinsic properties of the cells or a combination of both these factors influencing this age-related change, investigating changes in the BM cells are key to elucidating the mechanisms of these age-related changes. The fact that the stem cells residing within the BM are self-renewing, being used for various cell therapy applications and are also the progenitors of cells of different lineages and functions makes them the ideal candidates for these investigations and for exploring age-related changes at the single-cell and genetic levels.
2.2. Age-Related Changes in Haematopoietic Stem Cells (HSCs)
HSCs were the first identified stem cells and, to this date, remain the stem cells that have been explored extensively for their self-renewal abilities throughout the entire lifetime of an individual. However, they lose their ability to regenerate and replenish the other cell types rising from them with advancing age
[35]. While studies have indicated that their numbers increase in the elderly, it has been debated that this increase is a compensatory mechanism for their loss of functionality in the BM. Reports have also suggested that larger fractions of HSCs in the aged are less quiescent and undergo cell division as compared to those in the young, potentially due to the accumulation of higher levels of ROS over time
[36]. This, in turn, affects the repopulating capacity of HSCs in the elderly. Strzelecka and Damm discussed in depth the changes observed in aging haematopoietic cells, outlining that the readout of the transcriptome is a good indicator of the cellular state
[37].
Along with an increase in the number of cells with decreased functionality, the regenerative and homing capacities of aged HSCs are also shown to be lower in the BM. They suffer metabolic changes
[38], impaired autophagy and disrupted protein homeostasis, which altogether impact the regeneration potential of BM HSCs
[39]. Furthermore, the myeloid bias is enhanced during both inflammation, as well as during advancing age, leading to myeloid skewing in HSCs, indicating further responses towards the concept of inflammaging
[16]. In young individuals, haematopoiesis is normally supported via various HSC clones with similar potential. However, in the elderly, in spite of an overall increase in the number of HSCs, only a smaller proportion of the clones contribute to blood formation
[40]. This condition is known as clonal haematopoiesis of indeterminate potential or CHIP, which has increasingly been observed in people between the ages of 55 and 60 and has been linked with haematopoietic malignancies
[41].
2.3. Age-Related Changes in Mesenchymal Stem Cells (MSCs)
MSCs (also referred to as skeletal stem cells or SSCs) within the BM are the cells that can form into at least three types of tissues—the bone, fat and cartilage (property often referred to as trilineage differentiation). They are also known for their trophic factors and immunomodulating properties, making them very desirable for extensive use in cell therapy applications
[42]. These cells are usually identified in vitro by plastic adherence; trilineage differentiation and by the expression of CD73, CD105 and CD90 as per the International Society of Cell Therapy (ISCT)
[43]. However, other markers like SSEA4 and phenotypes CD45
low and CD271
+ are also used for the identification of uncultured BM MSCs
[44].
These cells are well-known for regenerative properties, which are reported to decline with advancing age. Studies have reported a decline in their number
[45][46], decline in their proliferative potential
[47][48], increased bias towards adipogenesis at the expense of osteoblast formation, resulting in decline in bone formation, and enhanced fat formation
[49] and decrease in migratory properties
[50]. More recently, potential links have been made between the intrinsic type 1 interferon profile within MSCs and their ability to respond to DNA damage
[51]. The work by Josephson et al. established the effect of age-related inflammation in MSCs by demonstrating a decline in the number of MSCs and impaired bone regeneration capacity in mice models. They also found that ‘circulating systemic factors’ in the BM of the aged contributed to MSC aging, confirmed by the presence of higher levels of TNF-α, IFN-γ and IL-6
[48], which are also classical indicators of inflammaging.
 +1 point
+1 point