1000/1000
Hot
Most Recent
 +1 point
+1 point
Cadmium (Cd), chromium (Cr) and lead (Pb) are heavy metals that have been classified as priority pollutants in aqueous environment while methane-oxidizing bacteria as a biofilter arguably consume up to 90% of the produced methane in the same aqueous environment before it escapes into the atmosphere. However, the underlying kinetics and active methane oxidizers are poorly understood for the hotspot of epipelon that provides a unique micro-ecosystem containing diversified guild of microorganisms including methane oxidizers for potential bioremediation of heavy metals. In the present study, the Pb2+, Cd2+and Cr6+ bioremediation potential of epipelon biofilm was assessed under both high (120,000 ppm) and near-atmospheric (6 ppm) methane concentrations. Epipelon biofilm demonstrated a high methane oxidation activity following microcosm incubation amended with a high concentration of methane, accompanied by the complete removal of 50 mg L−1 Pb2+ and 50 mg L−1 Cd2+ (14 days) and partial (20%) removal of 50 mg L−1 Cr6+ after 20 days. High methane dose stimulated a faster (144 h earlier) heavy metal removal rate compared to near-atmospheric methane concentrations. DNA-based stable isotope probing (DNA-SIP) following 13CH4 microcosm incubation revealed the growth and activity of different phylotypes of methanotrophs during the methane oxidation and heavy metal removal process. High throughput sequencing of 13C-labelled particulate methane monooxygenase gene pmoA and 16S rRNA genes revealed that the prevalent active methane oxidizers were type I affiliated methanotrophs, i.e., Methylobacter. Type II methanotrophs including Methylosinus and Methylocystis were also labeled only under high methane concentrations. These results suggest that epipelon biofilm can serve as an important micro-environment to alleviate both methane emission and the heavy metal contamination in aqueous ecosystems with constant high methane fluxes.
Heavy metals and the global rise in methane level both are serious threats to the environment and humans directly or indirectly [1][2]. According to the National Oceanic and Atmospheric Administration (NOAA) of USA, global methane emission has increased up to 1858 ppb till 2019, compared to pre-industrial times (722 ppb), and its impact on the environment has been 34 times greater than CO2 over a period of 100 years, as stated by the Environmental Protection Agency (EPA). It thus poses great challenges to address global warming and climate change by optimizing and developing the strategies for the reduction of the average level of global methane emissions, considering that it traps 21 times more heat than CO2 [3].
Wetland is an important target of methane reduction contributing approximately 20% of the global methane budget, while organic and inorganic sources of water pollution resulted in 1.8 million deaths during the year 2015 [4]. Heavy metals have emerged as a serious water contaminant in the present scenario as they tend to accumulate in the human body and result in mutagenic, teratogenic and carcinogenic changes[5][6]. According to the US-EPA, lead (Pb), cadmium (Cd) and chromium (Cr) are designated as the top priority contaminants that are of major public concern[7]. Numerous studies have been focused on physico-chemical approaches, such as ion exchange, flocculation, membrane filtration, biosorbents, electrodialysis, reverse osmosis and precipitation, for heavy metal removal in recent decades[8][9][10]. Though some of these techniques are effective in the removal of heavy metals, practical applications are not easily employed on a large scale due to their high cost and the secondary contaminations by the chemicals used in the processes. Several biological methods such as immobilization by biochar and rice straw, hyper accumulator plants, specific functional microbes have already been reported for heavy metals removal[11][12]. However, they require high labor cost and are not fully suitable in many cases under in situ conditions. Therefore, there is a need for a cost-efficient and environmentally sustainable technology that can remove heavy metals and methane gas simultaneously.
Methanotrophs have emerged as a tool of bioremediation due to the presence of methane monooxygenase (MMOs: pMMOs and sMMOs) enzymes with a distinctive capability of utilizing a vast array of compounds[13], including aliphatic halides and aromatic derivatives. The unique chemical reactivity and comprehensive substrate profiles of methane monooxygenases represent its potential implementation in bio-remediation of heavy metals[14].
In the last decade, a few studies have described the removal of heavy metals by methanotrophs. Methylosinus, a type II methanotrophs contributed 11–21% of the total bacterial DNA in the biofilm after addition of chromium (Cr) metal[15]. Additionally, the presence of Hg(II) and As(V) reductases was also confirmed in almost eight genera of methanotrophs, demonstrating that the metabolic potential of methanotrophs is generally overlooked[16]. Methylococcus capsulatus (Bath) was found to effectually bioremediate aquatic pollution caused by a broad range of chromium(VI) concentration (1.4–1000 mg L−1 of Cr+6)[17]. However, these methods took a longer time duration for the removal of heavy metals (20–90 days), moreover, the taxonomic identities of active methanotrophs remain poorly understood during the removal of heavy metals in complex aquatic environment.
Epipelon are periphytic biofilms that are known to promote the bio-stabilization of sediments and regularize benthic-pelagic nutrient cycling in benthic-pelagic zones of marine environments[18]. Periphyton are also known as an in situ strategy that has a promising role to stabilize micro-ecosystem, as well as in the restoration of devastated surface water ecologies[19] while the complex microbial communities and porous structures within can retain heavy metals[20]. Many microbial communities in periphytons are capable of adsorbing heavy metals by the secretion of extra polymeric substances (EPS) from cyanobacteria, green algae, diatoms and a variety of other bacteria[21]. Due to the intricate community structures, periphytic biofilms have a high resilience and capability to habituate to a wide range of concentrations of heavy metals and can maintain sustainable metabolic activities[22][23]. Periphytic raw material is also readily available and has a high metal removal and sorption potential, due to functional polar groups that are abundantly found on the periphytic surface[24], whereas, sludge after pollutant sorption, is less effective when compared to all other treatments. The presence of methanotrophs in lake sediments, as well as in wetland periphytons, has already been reported[25], but the methane oxidation capability of these biofilms have yet to be explored and the information regarding the capability of periphytic biofilms for simultaneous heavy metals removal and methane from aquatic ecosystems are scarce.
In the present study, a DNA-based stable isotope probing (DNA-SIP) technique was exploited to establish the correlation between the taxonomic identity of dynamic methanotrophs during heavy metal bioremediation and oxidization of methane in lake epipelon. The main objectives of the present study were: (1) to assess the heavy metals removal efficiency in association with increased methane oxidation capacity of epipelon; (2) to establish direct link between taxonomic identities of active methanotrophs and heavy metals bioremediation in epipelon; (3) to develop a novel strategy for mechanistic understanding of methanotroph-mediated feedback of microbial community structure for heavy metals bioremediation.
Epipelon biofilm was obtained, following standard methods, from the sediments of Xuan Wu, Lake Nanjing by using a 5-cm diameter acrylic core. The supernatant water was heedfully removed with a pipette to prevent planktonic algal contamination[26]. For further analysis, samples were collected from the top (1 cm) of the extruded sediment. The average conditions of the sampling site were determined at the time of sampling. The collected biofilm was transported and grown in the laboratory as previously described [19][27]. The epipelon biofilm was grown in 5 L glass beakers covered with perforated Sigma-Aldrich Parafilm®. The temperature was set to 28 °C and the beakers were placed in a growth chamber under a specific time period regime for periphytons (16 h light + 8 h dark period). A continuous 8 h dark and 16 h light period was used because epipelon biofilms proved to be light sensitive[28]. pH was adjusted to 6.80 for promotion of microbial growth as previously reported[19][27]. After 90 days of incubation, the biofilm was suspended and 5 mL of suspended biofilm, along with 15 mL of modified WC (without Carbon Source) was added to each microcosm and transferred into a 120 mL serum bottle as one microcosm. All microcosms were kept in a growth chamber for 20 days to stabilize epipelon growth. After the stabilization of biofilm, microcosms were further employed for the methane oxidation and heavy metal removal studies.
Microcosms for methane gas oxidation were prepared with the following six treatments in triplicate including controls. (1) 6 ppm 13CH4, (2) 120,000 ppm 13CH4 (12%), (3) 120,000 ppm 12CH4, (4) 50 mg L−1 Cr6+ + 120,000 ppm 13CH4, (5) 50 mg L−1 Pb2+ + 120,000 ppm 13CH4, and (6) 50 mg L−1 Cd2+ + 120,000 ppm 13CH4. The headspace gas of microcosm comprised of 67% N2 and 21% O2 for incubation conditions under 120,000 ppm (12%) methane concentration, or of 78% N2 and 21% O2 for incubation under near atmospheric methane condition (6 ppm). The stock solutions of metal salts, i.e., K2Cr2O7, PbCl2 and CdCl2 (each 500 mg L−1) were formed using deionized water.
To study the effect of methane oxidation on heavy metal bioremediation, six treatments with high and near atmospheric methane dose along with N2 and O2 were carried out as described earlier using microcosms in the laboratory: (1) 50 mg L−1 Cr6+ + 120,000 ppm 13CH4, (2) 50 mg L−1 Pb2+ + 120,000 ppm 13CH4, (3) 50 mg L−1 Cd2+ + 120,000 ppm 13CH4, (4) 50 mg L−1 Cr6+ + 6 ppm 13CH4, (5) 50 mg L−1 Pb2+ + 6 ppm 13CH4 and (6) 50 mg L−1 Cd2+ + 6 ppm 13CH4.
For each microcosm, 20 mL epipelon mixture was incubated in serum bottles sealed with a butyl stopper, after replacing head space volume with respective gases[29]. The 13CH4 used in the study was > 99% 13C-atom pure (Cambridge Isotope Laboratories, Tewksbury, MA, USA). Gas chromatography 2010-pro (Shimadzu GC12-A, Kyoto, Japan) was used to measure the concentration of the headspace CH4 every 24 h. Microcosm incubation was considered complete when the methane concentration was ≤100 ppm, and the epipelon biofilm was collected from each microcosm right after the treatment and all the samples were stored at −20 °C for further analysis. In order to determine the relative abundance of 13C-atom after microcosm incubation, substantially, 1 g sample was vacuum freeze-dried and was analyzed by a Flash 2000 elemental analyzer coupled to a Delta V™ IRMS Advantage isotope ratio mass spectrometer (Thermo Scientific™, Waltham, MA, USA). For heavy metals determination, a few drops of concentrated nitric acid were added to avoid precipitation of metals as well as growth of biofilm[30]. The samples were stored at 4 °C for further analysis. The concentrations of Pb2+, Cd2+ and Cr6+ in the samples were evaluated by inductively coupled plasma mass spectrometry (ICP-MS).
The morphology of the microorganism of epipelon biofilm was compared before and after treatment of 120,000 ppm 13CH4, by cell observation under SEM (EVO 18, Zeiss, Germany) and phase contrast microscopes (PCM) (Nikon Eclipse 135 Ti, Kyoto, Japan). The microbial activity for different substrates was assessed using the Biolog™ plate technique. Briefly, 2 g biofilm pellet was (centrifuged at 12,000 rpm and 25 °C) inoculated into plate wells[31] and was incubated at 25 °C for a week. The average well color development (AWCD) was monitored using a Biolog™ Microplate Reader ELx808 (590 nm) on a daily basis[27].
DNA was extracted from 500 mg epipelon sample of each treatment using the FastDNA spin kit for soil (MP Biomedicals, Santa Ana, CA, USA), according to the manufacturer’s manual. The quality and quantity of epipelonic DNA was evaluated using a NanoDrop ND-1000 UV-visible light spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). For 12CH4 and 13CH4 fractions, 13C-labeled DNA was isolated from 12C DNA by using density gradient centrifugation of total DNA, as formerly described[32]. The fractionated DNA in the CsCl medium was precipitated by polyethylene glycol 6000 (PEG 6000) and diluted to 30 µL water for downstream analysis.
Methanotrophic biomarker pmoA genes were quantified by real-time quantitative PCR (qPCR) from total DNA to ascertain the abundances of Methane Oxidizing Bacteria (MOB) on a CFX96 Optical Real-Time Detection System (Bio-Rad, Irvine, CA, USA). Fractionated DNA gradients of no. 2–14 were also used for the qPCR analysis of pmoA genes to assess the competence of 13C amalgamation into the genomic DNA of MOB communities. The primer pair and thermal cycling condition are given in details in the Supplementary Material (Table S1). PCR reaction mixtures and the standards were prepared and utilized as described previously[29]. Amplification efficiency ranged from 92% to 99%, with R2 values of 0.996 to 0.999.
Amplicon sequencing of 16S rRNA genes was conducted for the total DNA extracts, as well as DNA from the heavy CsCl fractions (buoyant density around 1.735 g mL−1) recovered from 13CH4-amended microcosms[33], using universal bacterial primer pair 515F/907R. Fractions with the identical buoyant density from 12CH4 microcosms was also sequenced to provide background details. Low-quality sequences were eliminated from generated reads by applying key-quality control steps[34] using QIIME pipeline[35]. A total of 696,738 sequences from 16S rRNA genes were obtained with quality score > 20, without mismatched primers and ambiguous bases (Table S2) and RDP MultiClassifier was used to obtain taxonomic assignments[36]. The relative abundance of type I methanotrophs was obtained as an aggregate of sequences associated to Methylobacter, Methylomonas, Methylomicrobium, Methylosoma and Methylococcus and type II methanotrophs included Methylosinus and Methylocystis sequences. All MOB-affiliated 16S rRNA gene sequences were collected and corporated into OTU at 97% identity threshold[37]. For 13C-labeled 16S rRNA genes, representative sequences of dominant OTUs (containing ≥ 2% of MOB-like 16S rRNA gene sequences in at least one of the selected samples) were applied for the phylogenetic tree analysis using the neighbor-joining method in latest MEGA-X version with bootstrapping value of 1000 replicates in all treatments[38].
Amplicon sequencing of pmoA genes was conducted on DNA from heavy fractions of 13CH4-amended microcosms using A189f/mb661r, using a similar strategy as that described previously (Cai et al., 2016). The generated pmoA gene reads were applied to bioinformatic processing key steps as previously described[33][39]. Molecular analysis was based on a total of 244,839 high-quality sequences (Table S3). For the major pmoA OTUs (containing ≥ 3% of pmoA gene sequences in at least one of the samples), an emblematic sequence was utilized for the phylogenetic analysis by associating with previously reported sequences from GenBank.
The Vegan package in R 3.4.3 (https://www.reproject.org/) was employed to compute the α-diversity indices (OTU richness, Chao1, Simpson index, Evenness index, and Shannon index) of epipelon biofilm samples.
Network visualization and modular analyses of 16S rRNA sequence results were obtained using Gephi (http://gephi.github.io/). The topological properties of the networks, including betweenness centrality, average degree, average path length, and modularity were also computed using Gephi for all treatments. The size of each node is directly proportional to the relative abundance of the bacterial community (log (n+1)).
The functional capacity of the epipelon biofilm samples was assessed by using the Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt) version 1.0.0[40]. A similarity threshold of 97% of all OTUs was used in QIIME for the prediction of PICRUSt and the Green genes database (13_5_release) was employed as a clustering reference while metagenomic functional profiles were prognosticated using the predict_metagenomes.py script.
The physicochemical properties at the sampling site of epipelon biofilm were: ammonia 0.66 mg L−1, nitrate 0.78 mg L−1, total phosphorus (TP) 0.23 mg L−1, total nitrogen (TN) 2.12 mg L−1 and pH 7.78. Epipelon are an integration of a wide variety of microorganisms, including algae, bacteria and protozoa, that collaboratively live in or on fine-grained substrata or sand. However, these biofilms under ambient methane concentration were predominantly composed of cyanobacteria and phytoplankton (Figure 1a). Moreover, after incubation under 120,000 ppm methane, bacterial species outnumbered cyanobacteria and phytoplankton as shown by PCM (Figure 1b). SEM showed that the structure of epipelon biofilm was intertwined bundles, with voids of algae, and diatoms, while bacteria attached to these were evident in these structures. The spaces and tunnels found within epipelonic biofilms are likely to provide physical space for bacterial attachment and metals sorption sites and are clearly visible in the SEM (Figure 1c,d)[41].
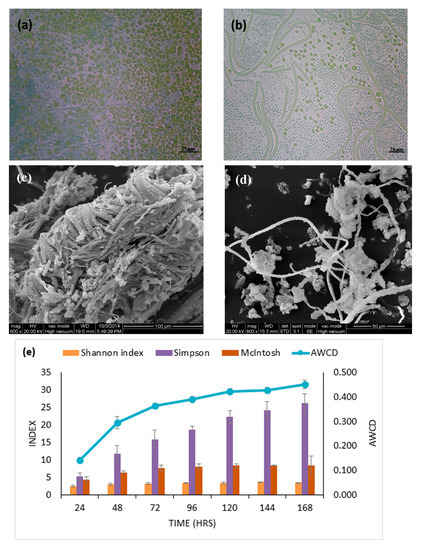
Figure 1. Phase Contrast Microscopy (PCM) and Scanning Electron microscopy (SEM) were performed to differentiate microbial community change (a) and (b) community composition of epipelon observed under PCM before and after 12% methane treatment, respectively. (c) and (d) structure of periphytic biofilm observed under SEM before and after 12% methane treatment, respectively. (e) The average well color development (AWCD) of the Biolog EcoPlates at 590 nm for periphytic biofilm and diversity indices (E) for species diversity.
The capability of different types of periphytic biofilms in the bioremediation and biosorption of different contaminants from physiochemically contrasting lakes has been intensively reported[42][43]. However, no studies have yet been reported about enhanced heavy metals removal by epipelon under a high methane regime which could likely represent naturally occurring conditions in a wetland.
In Biolog® experiments, the Average Well Color Development (AWCD) is an indicator of the degree of activity of epipelon biofilm. The darker color of the wells represents the higher activity of the microorganisms as confirmed from the slope of the graph (Figure 1e). The diversity indices showed higher community abundance, higher functional activity of epipelon, uniformity and species eveneness of the microbial community (Figure 1e), e.g., after incubation of Biolog® for 7 days, the Shannon index was found to be 3.6. This result indicates higher microbial diversity and species evenness in epipelon when compared to the control.
Epipelon was found to be highly active in removing Pb2+ and Cd2+ under 12% (120,000 ppm) methane, with 100% removal efficiency within 336 h (Figure 2a), although only 20% of Cr6+ was removed within same interval of time. It should also be noted that the low removal efficiency of Cr6+ could not entirely reflect the operating conditions in natural environments because Cr6+ is toxic in laboratory scale experiments without continuous recirculation, as previously suggested[44]. Meanwhile, hexavalent chromium is very toxic due to its high water-solubility and strong oxidizing ability. Chromium (VI) posed a toxicity risk to periphytic biofilm via inhibition of its photosynthetic activity and carbon utilization function[45], thus inducing destruction of the biofilm, which ultimately results in releasing Cr6+ back into the environment, making it more toxic. For instance, hexavalent chromium removal efficiency decreased from 90% to 50% for E. crassipes when compared to trivalent chromium[46]. A free water surface wetland made from sediment was found to be only capable of 7% chromium removal[47]. Furthermore, the isotope fractionation factor (ε) of Cr6+ was 2.62 ± 0.20% after 509 days of continuous incubation with methane based on theoretical estimation using the Rayleigh distillation mode[48] during slow bioremediation of Cr6+ by methanotrophs. In one similar periphytic study, Cd(II) was more secluded in comparison to Cr(VI), while field validation of this observation remained an intriguing issue for further research[49].
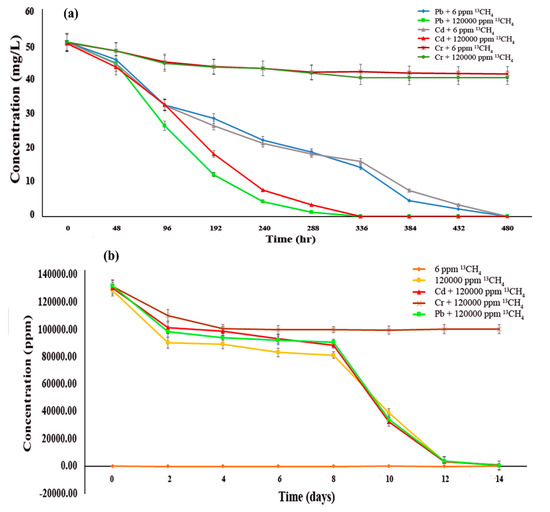
Figure 2. Methane oxidation and heavy metals bioremediation potential of epipelon (a) the ability of epipelonic biofilm to remove heavy metals at different time intervals coupled with methane oxidation (b) Methane oxidation at different time intervals under heavy metals effect.
It was further observed that under ambient methane concentration (6 ppm), epipelon completely removed amended Pb2+ or Cd2+ (both at 50 mg L−1) within 480 hrs, but the incubation under high methane concentration (120,000 ppm) greatly enhanced the metal removal efficiencies, which took only 336 hrs to complete the process (Figure 2a). This might be attributed to saturation effect of the active sites on the surface of the epipelonic biofilms. Comparatively, limited sensitivity of methane oxidation was found after adding Pb(II), which might have been due to soil water content as the primary key force for Pb(II) removal[50].
The patterns of metal tolerance, as shown by methane oxidizers, are comparatively distinct to chloride salts of Cd(II), Cr(III), Hg(II), Cu(II)and Zn(II)[51]. Another interesting aspect of the novel isolates of methanotrophs was their resistance to various types of pollutions (heavy metals, arsenate or organic) and were considered to be real ‘super-bugs’ due to their capability to withstand remarkably high concentration of various pollutants[52]. The abundance of extracellular polymeric substances, plays a key role in the higher removal efficacies of periphytic biofilms[21] as well as in-tank microbial activity[53].
The potential implication of methane monooxygenases, for bioremediation of different contaminants including heavy metals, is accentuated by their broad substrate profiles[54]. Hence, epipelonic biofilms have proven to be considerably effectual towards the removal of heavy metals within a shorter time period under high methane doses compared to previous studies, thus providing a reliable source for the removal of heavy metals from aquatic environments.
All treatments were found to exhibit strong methane oxidation activity and consumed more than 99% of amended methane except for 6 ppm 13CH4 treatment, which shows no significant methane oxidation, and 50 mg L−1 Cr6+ + 120,000 ppm 13CH4 only utilized 23.94% of available methane (Figure 2b). No significant difference in methane oxidation rate was observed between 12CH4 and 13CH4 treatment in microcosms with high concentration.
The assimilation of methane derived carbon obtained by methane oxidation was increased in organic carbon in high dose (12%) methane treatments. The percentage of 13C atom was significantly increased from 1.08% 13C atom in the near-atmospheric dose 13C methane (6 ppm) treatment and in high-dose 12C methane treatments to 16.24%, 12.14% and 13.78% 13C atoms in 120,000 ppm 13CH4, 50 mgL−1 Cd2+ + 120,000 ppm 13CH4 and 50 mgL−1 Pb2+ + 120,000 ppm 13CH4 treatments, respectively (Figure S1). These results confirm the potential of epipelonic biofilms for the assimilation of methane in higher amounts, thus confirming their potential towards effective methane oxidation in aquatic environments.
The abundance of the total microbiome and methanotrophs in epipelon was evaluated by real-time quantitative PCR of 16S rRNA and pmoA genes, respectively. Heavy metals significantly inhibit the abundance of total microbial communities (Figure 3d). Compared to control treatment (120,000 ppm 13CH4), the number of 16S rRNA genes was 37.94% and 26.49% lower in microcosm amended with Cd2 and Pb2+, respectively (Figure 3d). In previous studies, single cultures of methanotrophic bacteria were verified for bioremediation of heavy metals such as chromium[17][52][55]. Although the favorable conditions were revealed for a single type of strain, it is not a cost-effective and sustainable solution for heavy metal removal on a large scale. In the present study, the presence of Cr6+ in 50 mg L−1 Cr6+ + 120,000 ppm 13CH4 treatment resulted in a lower abundance (6.97 less folds) of methanotrophs due to Cr6+ toxicity as compared to 120,000 ppm 13CH4 whereas the treatments with lead (1.20 less folds) and cadmium (1.57 less folds) did not show much effect on the methanotrophic community in biofilms.
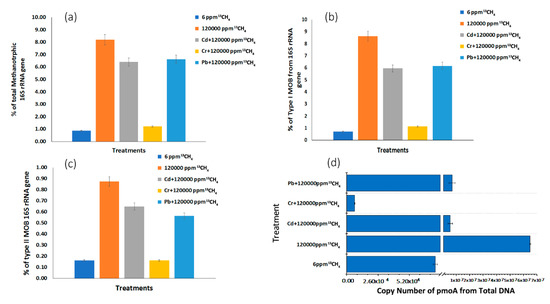
Figure 3. Illumina sequencing was carried out at the whole epipelon microbial community level in microcosms targeting total methanotrophs from total 16S rRNA genes (a), and the relative abundances of type I (b), type II (c), The pmoA gene copy numbers were assessed using real-time quantitative PCR (d).
On the other hand, 120,000 ppm 13CH4 got 884, 12.13 and 9.46 fold higher MOB than 6 ppm 13CH4, 50 mg L−1 Cd2++ 120,000 ppm 13CH4, 50 mg L−1 Pb2++ 120,000 ppm 13CH4, respectively (Figure 3d).
According to high-throughput sequencing of 16S rRNA genes, high methane treatments (120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 and 50 mg L−1 Pb2++ 120,000 ppm 13CH4 ) got 9.7, 7.35, 7.69 more folds of MOB genes, respectively, compared to near-atmospheric methane treatment (6 ppm 13CH4) (Figure 3a). Chromium-augmented treatment showed only 1.41 more folds of MOB genes than near atmospheric methane treatment. This might be due to toxic nature of chromium that suppressed the abundance of MOB genes in the following treatments.
The abundance of type I and type II MOB in the whole microbial community was determined using high-throughput sequencing of 16S rRNA genes data. Type I methanotrophs copy numbers were increased by factors of 12.16, 8.39, 8.69 fold in the high methane treatments (120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 and 50 mg L−1 Pb2+ + 120,000 ppm 13CH4) compared to near-atmospheric methane treatments (6 ppm) (Figure 3b). Type II MOB was also stimulated by 5.43, 4.02, 3.50 and 1.01 folds in 120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4, 50 mg L−1 Pb2+ + 120,000 ppm 13CH4 and 50 mg L−1 Cr + 120,000 ppm 13CH4, respectively, as compared to 6 ppm 13CH4 (Figure 3c).
The chromium-augmented treatment showed only a 1.61-fold increase for the high methane as compared to near atmospheric methane treatment (6 ppm). The sequence analysis of the microbial community from lake Biwa, Japan [3][56], stratified eutrophic Swiss lakes[57] and Lacamas lake in Washington, USA[58] showed Type I methanotroph dominances, similarly to our findings in this study.
The heavy fraction resolved by isopycnic centrifugation of the DNA was employed for qPCR of pmoA genes and clearly indicates active cell propagation during the consumption of 13C-methane in all the high methane dose treatments, except for the Cr6+ amended (10.05%) treatment and ambient 13CH4 (6 ppm) treatment. A peak shift of relative pmoA gene abundances (Figure 4a) towards heavy fractions (buoyant density around 1.745 g mL−1) was very evident in all high-dose 13CH4 treatments, while the 12CH4 pmoA gene abundances peaked only in the light fractions (buoyant density around 1.72 g mL−1)[29]. In 13C-labeled heavy DNA fractions (buoyant density around 1.745 g mL−1), the number of pmoA genes was significantly increased to 57.73%, 61.79% and 62.62% in 120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 and 50 mg L−1 Pb2+ + 120,000 ppm 13CH4, respectively (Figure 4b) as compared to the non-significant 13CH4 (6 ppm) treatment.
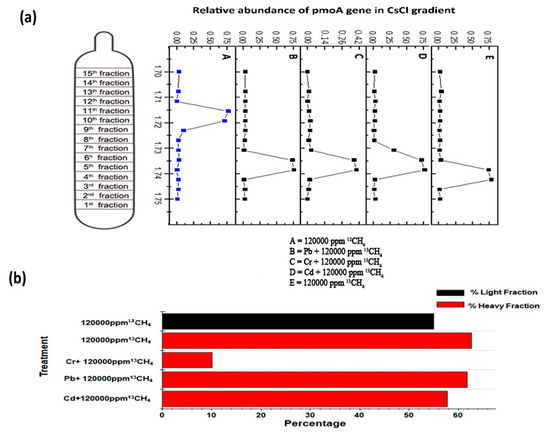
Figure 4. The enrichment of 13Clabeled methanotrophs on the basis of qPCR of pmoA gene (a). The relative pmoA gene abundance was the proportion of MOB like gene copy numbers in each fraction to the total abundance through the gradient in a treatment (b). The red columns represent the percentage of MOB affiliated reads in total 16S rRNA genes in the heavy fractions (buoyant density 1.745 g mL−1) while the black column represents light fraction (buoyant density 1.72 g mL−1).
The structures and abundances of microbial communities were drastically changed due to the selection of distinct phylotypes in the different heavy metal treatments, along with methane oxidation (Figure S2). The taxonomic analysis of heavy metal resistant genes revealed that the most common bacterial phyla in the treatments was Proteobacteria followed by Actinobacteria[59] which is in consistent with our study. Microbial community composition and interaction was analyzed by high throughput sequencing to determine the impact of metals and methane oxidation on microbial community structure. The taxonomic classification of OTUs at different levels produced 28 phyla, 92 classes, 178 orders, 334 families and 751 genera. Approximately 28 different phyla were found to be present in sequencing of the epipelonic biofilm samples, among these Proteobacteria, Cyanobacteria, Saccharibacteria and Gemmatimonadetes were the most abundant phyla in all treatments (Figure S3). The phylum Proteobacteria was found to be the most dominant in 120,000 ppm 13CH4. In comparison to other treatments, it outnumbered 50 mg L−1 Pb2+ +120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 and 6 ppm 13CH4 by 13.66%, 17.23% and 22%, respectively. Alpha and gamma Proteobacteria have recently been shown to be effectual for the bioremediation of different heavy metals such as cadmium in an iron reducing bacterial immobilization system[60].
On the basis of Spearman correlation analysis, a co-occurrence network of different co-existing communities 120,000 ppm 13CH4 comprised of 287 nodes, 5128 edges, average degree 1.87, highest graph density 0.182, highest modularity 0.193 and average network distance 2.58 was generated, indicating strong microbial interactions (Figure 5). Ambient 6 ppm 13CH4 and 50 mg L−1 Cr6+ + 120,000 ppm 13CH4 treatments were dominated by alpha-Proteobacteria but the scenario was changed to gamma Proteobacteria in high methane dose treatments as represented by colored dots in co-occurrence network. Based on the level of interactions from all the calculated modules, we can arrange treatments in the following decreasing order of interaction: 120,000 ppm 13CH4 > 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 > 50 mg L−1 Pb2+ + 120,000 ppm 13CH4 > 6 ppm 13CH4 > 50 mg L−1 Cr6+ + 120,000 ppm 13CH4 (Table S4). The modularity of all the networks differs from each other, demonstrating the verity that the modularity of different networks with different doses of methane or metals or contaminants changed[61]. However, the strong correlation between nodes illustrates that the microbe–microbe interaction stayed strong after the intrusion of external sources or by the bioadsorption of heavy metals or methane oxidation.
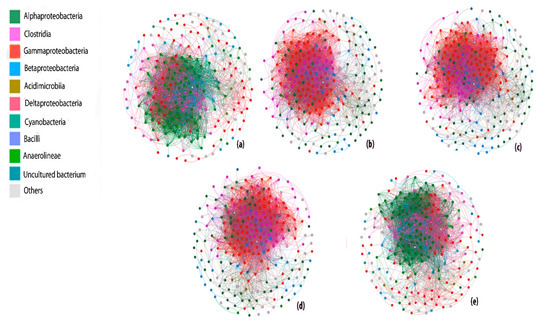
Figure 5. The co-occurrence network analysis of bacterial communities in the order (a) 6 ppm 13CH4 (b) 120,000 ppm 13CH4 (c) 50 mg L−1 Pb2+ + 120,000 ppm 13CH4 (d) 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 (e) 50 mg L−1 Cr6+ + 120,000 ppm 13CH4. The network represents strong (Spearman’s r > 0.8) and significant (p value < 0.01) correlation. Each node represents a unique sequence in the whole data set, colored nodes (excluding light gray) depicts the ten major groups while light gray represents all other groups.
Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt) analysis represented functional attributes of all treatments, 120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 and 50 mg L−1 Pb2+ + 120,000 ppm 13CH4 was dominated by methanotrophy (Figure S4) followed by chemohetrotrophy, aerobic chemohetrotrophy, and phototrophy. Pheatmap confirms the presence of genes engaged in numerous processes such as cellular processes, environmental information processing, metabolism, and genetic information processing[62].
We further investigated the presence of dominant species at phylum, class, order, and family level by linear discriminant analysis effect size pipeline (LEfSe) analysis. Proteobacteria dominated among all treatments while the 50 mg L−1 Cr6+ + 120,000 ppm 13CH4 community was dominated by Cyanobacteria and Acidobacteria (Figure S5). The microbial community in treatments, 50 mg L−1 Cr6+ + 120,000 ppm 13CH4 was predominately composed of Verrucomicrobia, Methylomonas and Fimbriimonadales.
Taxonomic analysis of 13C-labeled methanotrophic 16S rRNA genes further demonstrated distinct proportions of active type I and II methane oxidizers in all treatments (Figure 6b). Particularly, Methylobacter a type I methanotroph was 51.81%, 48.09%, 49.59% abundant in 50 mg L−1 Pb2+ + 120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4, 120,000 ppm 13CH4 treatments, respectively (Figure 6a). On the contrary, Methylomicrobium was 80.04% abundant in 50 mg L−1 Cr6+ + 120,000 ppm 13CH4. These results were further confirmed by neighbor-joining tree-clustering around Methylobacter spp. (Figure 6c).
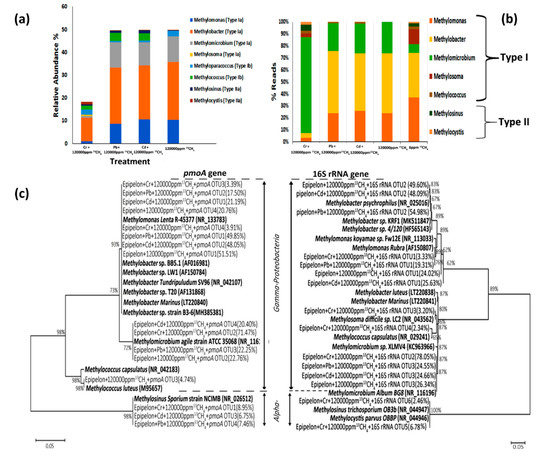
Figure 6. Methanotrophic community compositions of 13C-labeled MOB based on phylogenetic tree analysis of 16S rRNA and pmoA genes. (a) Percentage of type Ia, Ib, II and IIa methanotrophs were computed from the 13C-labeled genes. (b) Percentages of type I and II MOB were determined from16S rRNA high throughput sequencing of the 13C-labeled genes (c) Neighbor-joining phylogenetic tree depicts the phylogenetic relationship of dominant OTU sequences with the known MOB sequences from GenBank. The percentages illustrate the relative sequence abundance of each OTU to the total MOB-affiliated 16S rRNA or pmoA genes. Bootstrap values higher than 60% are pointed at the branch nodes.
Thus, it was demonstrated that the abundance of Methylobacter sp. by the 13C-labeled pmoA genes. Type I outnumbered Type II with percentages of 90.65%, 90.91% and 99.80% of total methanotrophs in 50 mg L−1 Pb2+ + 120,000 ppm 13CH4, 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 and 120,000 ppm 13CH4, respectively. Type II MOBs, Methylosinus and Methylocystis, were stimulated by high methane dose and metal contamination in treatments 50 mg L−1 Pb2+ + 120,000 ppm 13CH4 and 50 mg L−1 Cd2+ + 120,000 ppm 13CH4 by 9.14% and 8.8% as compared to 120,000 ppm 13CH4.
Methylobacter spp. and Rhodoferax spp. were found to be the most dominant species in heavy metals contaminated landfill[63]. Zolgharnein, et al.[64] have evaluated the role of Delfetia and Methylobacter spp. in the removal of heavy metals such as lead, zinc, copper and cadmium, for the first time. They reported that in the absence of Cu(II) or Cu(I), methanobactin from Methylosinus trichosporium OB3b will bind Ag(I), Co(II), Au(III), Cd(II), Hg(II), Fe(III), Ni(II), Mn(II), Pb(II), Zn(II), or U(VI) which represents the ability of Methylosinus to bioremediate heavy metals[55]. Similarly, Methylococcus capsulatus has been found to detoxify lead and zinc at low pH [65] and chromium[17]. The type II methanotrophic strains identified in this study have been widely utilized for the bioremediation of heavy metals[16][66].
These results imply that many methanotrophs, including Methylobacter, Methylococcus, Methylosinus, Methylocystis and Methylmicrobium have an unacknowledged metabolic potential and multifaceted capabilities for bio-remediating heavy metals. Higher type I methanotrophic activities were confirmed in all treatments, which might be due to the higher impact of intrinsic biotic and abiotic characteristics of Xuan Wu Lake.
In the present study, the potential capability of naturally occurring epipelon biofilms for the simultaneous removal of heavy metals along with methane oxidation was assessed. The results show that high methane dose (12%), has a substantial effect on heavy metals (Pb, Cd, Cr) removal. Epipelon exhibits a strong methane oxidation potential and 100% removal efficiency was achieved for Pb2+ and Cd2+ within 336 h, whereas Cr6+ was only removed by as little as 20% even after 480 h. Under high methane concentration (120,000 ppmv), bioremediation capability of epipelon is enhanced in terms of time. MOBs along with other co-existing microbes are dominated by type I after treating with 120,000 ppm (12%) headspace methane. The addition of Cr6+ has a negative impact on the epipelon community structure and inhibits methanotrophs along with other species. Heavy metals cause reduced abundance of certain microbes as compared to control treatments with 12% methane in the absence of metals. It is concluded from the present study that epipelon have strong potential to be used as an environmentally friendly technology which can be used for the simultaneous removal of different types of pollutants in aqueous and atmospheric environments.
The following are available online at https://www.mdpi.com/2076-2607/8/4/505/s1, Table S1. Primers and PCR conditions used in this study. Table S2. Summary of MiSeq amplicon sequencing of 16S rRNA genes from total DNA sample. Table S3. Summary of Illumina MiSeq amplicon sequencing of pmoA genes from total DNA. Table S4 Topological properties of the co-occurrence networks of lake microbial communities. Figure S1. Percentage of 13C atoms abundance accumulated by methanotrophs in Epipelon. Figure S2. The changes in community composition of microbes following microcosm incubation with respective methane doses based on high throughput sequencing of 16S rRNA genes at class level. Figure S3. The changes in community composition of microbes following microcosm incubation with respective methane doses based on high throughput sequencing of 16S rRNA genes at phylum level. Figure S4. Functional pheat map of Epipelon under different methane and metals doses. Figure S5. linear discriminant analysis effect size pipeline (LEfSe) analysis for dominant phyla, class and species.





