1000/1000
Hot
Most Recent
 +1 point
+1 point
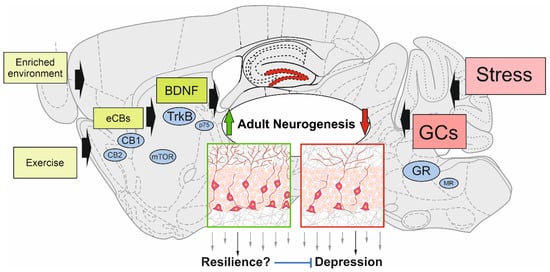
Newborn neurons in the adult hippocampus are regulated by many intrinsic and extrinsic cues. It is well accepted that elevated glucocorticoid levels lead to downregulation of adult neurogenesis, which this review discusses as one reason why psychiatric diseases, such as major depression, develop after long-term stress exposure. In reverse, adult neurogenesis has been suggested to protect against stress-induced major depression, and hence, could serve as a resilience mechanism.
| Protocol of Stress | Behavior | Effect on Neurogenesis (↓ Decreased; ↔ Unchanged; ↑ Increased) |
|---|---|---|
| Chronic social stress, Chronic social defeat stress (CSDS) |
↑ Anhedonia, Social avoidance, ↑ Sleep disturbances, ↓ Exploratory anxiety, ↓ Weight | ↓ Simon et al., 2005; Schloesser et al., 2010; Jiang et al., 2019 [52][53][54] ↔ Hanson et al., 2011 [55] ↑ Lagace et al., 2010 [56] |
| Chronic unpredictable stress (CUS), (Unpredictable) Chronic mild stress ((U)CMS) |
↑ Anhedonia, ↑ Sleep disturbances, ↑ Behavioral despair, ↓ Grooming, ↓ Weight | ↓ Jayatissa et al., 2006, 2009; Toth et al., 2008; Surget et al., 2011; Dioli et al., 2017 [57][58][59][60][61] ↔ Lee et al., 2006 [62] |
| Chronic corticosteroid treatment | ↑ Anhedonia, ↑ Behavioral despair, ↑ Anxiety | ↓ Ekstrand et al., 2008; Brummelte and Galea, 2010; Pazini et al., 2017; Levone et al., 2020 [63][64][65][66] |
| Repeated restraint stress | ↑ Anhedonia, ↑ Anxiety, ↑ Behavioral despair | ↓ Luo et al., 2005; Rosenbrock et al., 2005; Snyder et al., 2011 [38][67][68] ↔ O’Leary et al., 2012 [69] ↑ Parihar et al., 2011 [70] |
| Early life stress (ELS) | ↑ Anhedonia, ↑ Anxiety, ↑ Behavioral despair, ↓ Learning, ↓ Locomotion | ↓ Mirescu et al., 2004; Kikusui et al., 2009; Lajud et al., 2012 [71][72][73] |
| Prenatal (restraint of pregnant dams) | ↑ Anhedonia, ↑ Anxiety, ↑ Behavioral despair | ↓ Lemaire et al., 2000; Bosch et al., 2006; Mandyam et al., 2008; Lucassen et al., 2009 [74][75][76][77] |
| Learned helplessness (chronic tail or footshocks) (LH) | ↓ Active avoidance, ↑ Sleep disturbances, ↓ Weight | ↓ Malberg and Duman, 2003 [78] ↔ Van der Borght et al., 2005 [79] |
| Social isolation (SI) | ↑ Anxiety, ↑ Behavioral despair, ↓ Learning, |
↓ Westenbroek et al., 2004; Spritzer et al., 2011; Chan et al., 2017 [80][81][82] |
| Lipopolysaccharide-induced sickness behavior | ↑ Anhedonia, ↑ Lethargy, ↓ Appetite and food intake, ↑ Anxiety | ↓ Ekdahl et al., 2003; Monje, 2003; Yirmiya and Goshen, 2011; Perez-Dominguez et al., 2019 [83][84][85][86] ↔ Depino, 2015 [87] |