1000/1000
Hot
Most Recent
 +1 point
+1 point
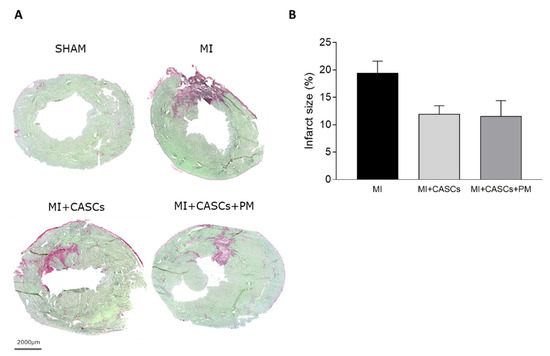
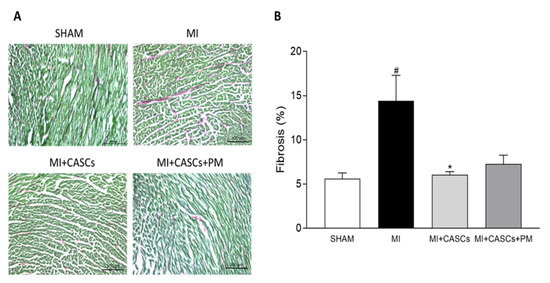
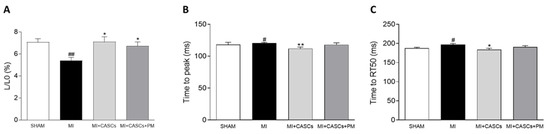
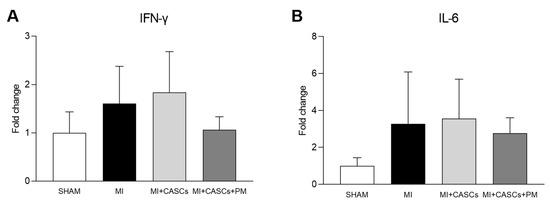
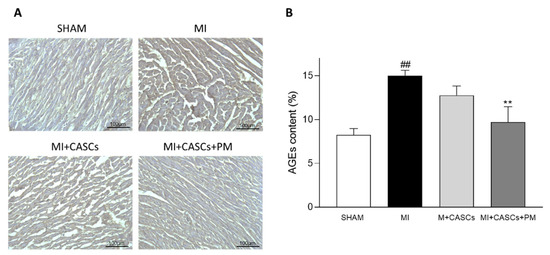
Myocardial infarction (MI) occurs when the coronary blood supply is interrupted. As a consequence, cardiomyocytes are irreversibly damaged and lost. Unfortunately, current therapies for MI are unable to prevent progression towards heart failure. As the renewal rate of cardiomyocytes is minimal, the optimal treatment should achieve effective cardiac regeneration, possibly with stem cells transplantation. In that context, our research group identified the cardiac atrial appendage stem cells (CASCs) as a new cellular therapy. However, CASCs are transplanted into a hostile environment, with elevated levels of advanced glycation end products (AGEs), which may affect their regenerative potential. In this study, we hypothesize that pyridoxamine (PM), a vitamin B6 derivative, could further enhance the regenerative capacities of CASCs transplanted after MI by reducing AGEs’ formation. Methods and Results: MI was induced in rats by ligation of the left anterior descending artery. Animals were assigned to either no therapy (MI), CASCs transplantation (MI + CASCs), or CASCs transplantation supplemented with PM treatment (MI + CASCs + PM). Four weeks post-surgery, global cardiac function and infarct size were improved upon CASCs transplantation. Interstitial collagen deposition, evaluated on cryosections, was decreased in the MI animals transplanted with CASCs. Contractile properties of resident left ventricular cardiomyocytes were assessed by unloaded cell shortening. CASCs transplantation prevented cardiomyocyte shortening deterioration. Even if PM significantly reduced cardiac levels of AGEs, cardiac outcome was not further improved. Conclusion: Limiting AGEs’ formation with PM during an ischemic injury in vivo did not further enhance the improved cardiac phenotype obtained with CASCs transplantation. Whether AGEs play an important deleterious role in the setting of stem cell therapy after MI warrants further examination.
| Parameters | 4 Weeks Post-Operative | |||
|---|---|---|---|---|
| SHAM | MI | MI + CASCs | MI + CASCs + PM | |
| EF (%) | 80 ± 4 | 59 ± 4 ## | 79 ± 3 *** | 72 ± 3 |
| HR (bpm) | 333 ± 14 | 318 ± 12 | 332 ± 12 | 327 ± 13 |
| SV (µL) | 170 ± 16 | 162 ± 17 | 172 ± 17 | 192 ± 17 |
| CO (mL/min) | 57 ± 4 | 51 ± 5 | 59 ± 6 | 65 ± 6 |
| EDV (µL) | 215 ± 26 | 298 ± 51 | 217 ± 17 | 267 ± 22 |
| ESV (µL) | 45 ± 13 | 136 ± 36 | 44 ± 5 * | 77 ± 11 |
| AWT (mm) | 1.74 ± 0.13 | 1.55 ± 0.16 | 1.65 ± 0.12 | 1.80 ± 0.16 |
| PWT (mm) | 1.54 ± 0.16 | 1.63 ± 0.15 | 1.57 ± 0.09 | 1.62 ± 0.17 |
| Parameters | 4 Weeks Post-Operative | |||
|---|---|---|---|---|
| SHAM | MI | MI + CASCs | MI + CASCs + PM | |
| Max LV pressure (mmHg) | 99 ± 3 | 90 ± 2 | 96 ± 3 | 103 ± 4 * |
| dP/dtmax (mmHg/s) | 6773 ± 529 | 6038 ± 242 | 6923 ± 340 | 6551 ± 257 |
| dP/dtmin (mmHg/s) | −7269 ± 683 | −6550 ± 705 | −6816 ± 354 | −6917 ± 273 |
| Tau (s) | 0.0130 ± 0.001 | 0.0499 ± 0.017 | 0.0148 ± 0.002 | 0.0117 ± 0.001 ** |