Escherichia coli Nissle 1917 (EcN) is a commonly used probiotic in clinical practice. Its facultative anaerobic property drives it to selectively colonize in the hypoxic area of the tumor for survival and reproduction. EcN can be engineered as a bacteria-based microrobot for molecular imaging, drug delivery, and gene delivery.
1. Characteristics of EcN
EcN is a facultative anaerobic organism that proliferates mainly in the interface between the necrotic and hypoxic regions of tumors [1] and exists in rich oxygen areas [2], expanding their potential applications for various tumor types. Moreover, the special serum-sensitive lipopolysaccharide on the membrane of EcN promotes quick clearance from normal organs [3]. Researchers have systematically studied the biodistribution and quantitative tumor colonization of EcN in vivo [4]. It was found that the tumor/liver ratio of EcN colonization after intravenous injection was at least 10,000:1 in 4T1 tumor-bearing BALB/c mice, giving EcN a massive edge over traditional nanomedicine in terms of tumor accumulation and overall biodistribution profile. Interestingly, the average number of EcN found in tumors was significantly higher than the injected dose due to the colonization and proliferation of EcN. The minimum bacterial dose for successful colonization was 20,000 CFU, at which the average number in the tumor reached 10 8 CFU. As the injection dose increases, the number of bacteria colonized in the tumor increases, but the bacteria in the liver and spleen grow accordingly. Moreover, the route of EcN injection, such as intravenous (i.v.), intraperitoneal (i.p.), and intertumoral (i.t.) injection, did not influence the tumor targeting and tumor-to-organ ratios. Oral administration of EcN confirmed that the bacteria crossed the gastrointestinal tract and colonized hepatic metastases [5]. Therefore, preferential tumor colonization of EcN may allow for flexible administration choices to meet specific clinical needs.
EcN has multiple peritrichous flagella that may drive it forward as a bio-engine [6]. Therefore, EcN can be developed into a self-propelled microrobot to break through the biological or pathological barriers to deliver therapeutic payloads [7]. The whole-genome sequencing of EcN has been completed [8], and methods for genetic modification of genomes and transformation of plasmid have been established to engineer EcN for heterologous gene expression [9][10][11]. Therefore, the therapeutic payloads can be drugs, expressed proteins, antigens, and immunoregulatory factors. However, constitutive expression of therapeutic factors may cause undesirable adverse effects, such as hepatic and splenic injury, so it is necessary to control the heterologous gene expression temporally and quantitatively. Weiss’s group established an in vivo remote control (IVRC) system to deliberate the external control of gene expression [12]. Three inducible promoter systems enabled EcN to remotely control and precisely regulate the kinetics of gene expression, and the L-arabinose–ParaBAD system showed the highest induce efficiency [13]. After oral administration or intraperitoneal injection of inducer L-arabinose, the expression of reporter gene luciferase in EcN colonized tumor reached its maximum after 6 h and stopped when L-arabinose was removed. Therefore, the controllable expression of EcN may provide a highly flexible and suitable treatment for individualized therapy. Precise regulation of the ECN number to control its proliferation in the tumor and expression of therapeutic agents will be of great significance to achieve spatiotemporal and quantitative imaging and treatment response. However, the underlying mechanisms of tumor targeting, and colonization of bacteria are complex and remain unclear. The influential factors may include the bacterial species used, types of tumor treated, and the tumor microenvironment [14]. Therefore, regulating the expression of bacteria may be a more practical means.
2. EcN-Mediated Tumor Imaging
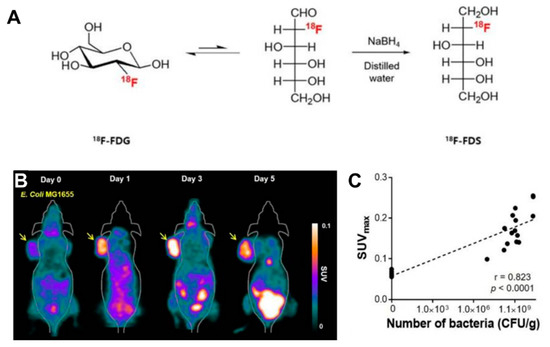
Radionuclide-based molecular imaging, namely, PET and SPECT, is a powerful tool to assess physiological and pathological processes in vivo without penetration depth limitation. Currently, radiopharmaceuticals for bacterial imaging focus on tracking bacteria to differentiate sterile inflammation from infection [15]. Based on the tumor-specific colonization nature of EcN, radiopharmaceuticals monitoring EcN can be used for tumor imaging. The living body itself contains various background bacteria, so radiotracers must be highly specific to target injected EcN. The endogenous bacterial thymidine kinase gene (TK gene) of EcN has been shown to be an effective reporter gene for nuclear medicine imaging using radiolabeled pyrimidine nucleoside analogs, such as [18F]-FEAU, [124I]-FIAU, and [125I]-FIAU [16][17]. Since the substrate of bacterial TK presents poorly binding affinity with mammalian TK, the radiotracers mentioned above can selectively identify and locate bacteria in vivo [18]. PET Imaging with [18F]-FEAU exhibited high accumulation in tumors and a linear correlation with the number of colonized EcNs, offering precise information about the survival, proliferation, and number of the bacteria. A strategy of engineered EcN with exogenous reporter genes hSSTR2 has been reported for in vivo tumor visualization [19]. The outer membrane protein receptor FyuA of EcN can selectively recognize the 64Cu and 89Zr labeled metallophore yersiniabactin (YbT), which has a high affinity for transition metals [20]. A substantially higher PET signal was also observed in the EcN colonized tumor than that without the bacterial injection. PET tracers targeting bacteria-specific sugar metabolism have also been developed, and [18F]-FDS is the most representative one. [18F]-FDS, a synthetic analog of [18F]-FDG, has been shown to accumulate in Gram-negative Enterobacteriaceae selectively but not in mammalian or cancer cells. In PET imaging, the uptake of radioactivity in the tumor had a positive relationship with the number of viable bacteria, allowing a semiquantitative measure of bacterial density in the tumor [21] ( Figure 1 ). The successful visualization and quantification of therapeutic E. coli by [18F]-FDS will make it possible to predict the therapeutic response, which could facilitate the clinical translation of bacteria-mediated tumor therapy.
Figure 1. (A) [18F]-FDS PET imaging in CT26-bearing mice treated with E. coli. (A) PET imaging was performed at day 0, 1, 3, 5 after intravenous injection of E. coli. The radioactivity uptake of the tumor was significantly higher at day 1, 3, 5 than in pre-treatment. (B) Positive correlation between SUVmax and the number of viable bacteria. (C) Schematic illustration of the synthesis of [18F]-FDS from [18F]-FDG. Reproduced with permission from Jung-Joon Min, Theranostics; published by Ivyspring International Publisher, 2020.
3. EcN-Mediated Tumor Therapy
The specific tumor targeting of EcN facilitates the establishment of a live platform for the delivery of cancer therapeutics. Three main strategies have been employed to achieve EcN-based tumor treatment: (1) to load the drug or nanoparticles as a microrobot for specific tumor delivery; (2) to engineer the EcN to express anticancer proteins for tumor management, and (3) to deliver the immuno-regulatory agent for cancer immunotherapy.
3.1. Direct Drug Delivery
For cancer treatment, the concentration of drugs in the tumor plays a key role in regulating the therapeutic effect. The EcN vehicles with self-propulsion ability may enhance the drug accumulation in tumor sites compared with passive drug diffusion. Once it reaches the tumor tissue, EcN drives itself to swim against the barriers of the tumor microenvironment and seek the hypoxic regions for colonization. Therefore, EcN-mediated drug delivery would penetrate the depth of the tumor to improve the antitumor efficacy. Doxorubicin (DOX) has been conjugated to EcN via acid-labile linkers, realizing a high DOX concentration in the tumor at the uptake value of ~12.9% of the injected dose per gram tissue (%ID/g) after 3 h of intravenous injection, which is much higher than the conventional nanocarriers
[2].
However, nanomaterials have the advantages of multi-functionalization and modification. The integration of bacteria with nanomaterials offers a new combinational and synergistic therapeutic approach to reduce their respective limitations and achieve complementary advantages. An amphiphilic copolymer PM
TOS/PM
DOX was obtained by conjugating poly(ethylene glycol) with doxorubicin (DOX) or α-tocopheryl succinate (TOS), and then immobilizing it onto the EcN through acid-labile linkers (namely EcN-PM
D/T). In response to the acidic environment of the tumor, copolymers were released from EcN and self-assembled into hybrid micelles (M
D/T) in situ. Then the GSH releases TOS and DOX to achieve tumor suppression. In treatment with free EcN, DOX/TOS mixtures, M
D/T hybrid micelles, and EcN-PM
D/T, the EcN-PM
D/T conjugates exhibited enhanced tumor growth inhibition with a longer survival rate
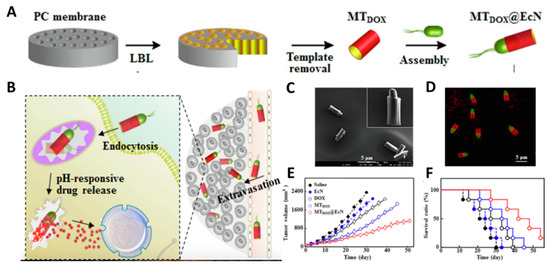
[22]. EcN can be successfully embedded into microtubes (MT
dox@EcN) as a biorocket
[23], which was confirmed by confocal laser scanning microscope (CLSM) images (
Figure 2). The motion of EcN enhanced the extravasation of MT
dox@EcN from the blood vessel, resulting in the high accumulation of DOX in the tumor. As shown in
Figure 2E, the tumor inhibition ratio of MT
dox@EcN (75.6%) is remarkably more prominent in vivo compared to EcN(20.6%), free DOX(36.1%), and MT
dox(60.9%). Furthermore, MT
dox@EcN exhibited a higher survival rate with a median survival time of 42 days, demonstrating stronger antitumor efficiency than other treatments. However, the microtubes also restricted the flagella movement, resulting in the lower velocity of MT
dox@EcN (6.8 μm/s) than that of free EcN (9.8 μm/s)
[2]. Reduction in motion ability directly influenced tumor targeting and penetration efficiency, which may be one reason MT
dox@EcN did not eliminate tumor tissues in the antitumor experiment. The balance between the loading of nano cargos and the mobility of bacteria should be considered in the construction of the bacteria-nanomaterials system. In general, the integration of drug-carrying micro/nanomaterials with bacteria will enable them to work synergistically to achieve advanced antitumor effects.
Figure 2. (A) Schematic illustration of the synthesis of MTdox@EcN; (B) Schematic illustration of the mechanism of MTDOX@EcN as a biorocket for drug delivery in tumor; (C) Typical SEM and (D) CLSM images of MTDOX@EcN. (E) Tumor inhibition and (F) survival rates of MTdox@EcN treatment in tumor-bearing mice. Reproduced with permission from Xiaohong Li, Chemical Engineering Journal; published by Elsevier, 2020.
To avoid the insecure factors caused by live bacteria and meet diverse medical needs, EcN can be prepared as minicells and bacterial ghosts (BGs). Minicells are the nanosized forms of bacteria that retain the same cytoplasmic components but lose the ability to proliferate due to genome deficiency
[24]. BGs are the nonliving membranes shells of bacteria without cytoplasmic and DNA content
[25]. Minicells and bacterial ghosts still retain the same tumor-targeting properties as their parent bacteria. The EcN-derived minicells displaying pHLIP could be used for delivering DOX and successfully invade the necrotic and hypoxic regions of orthotopic breast cancer
[26]. EcN BGs were reported to be foreign antigen carriers and drug carriers for treating eye diseases
[27].
3.2. Gene Therapy
EcN is a programmable vehicle designed to carry exogenous genes via Red/ET recombination or CRISP-Cas9 to express therapeutic proteins. EcN successfully expressed Tum-5, a powerful angiogenesis inhibitor. In mice bearing B16F10 mouse melanoma tumors, EcN (Tum-5) demonstrated remarkable tumor suppression after the upregulation of Tum-5 expression
[28]. Then the Tum-5-p53 bifunctional proteins were constructed and engineered into EcN
[29]. The decreased tumor volume and tumor weight indicated that the antitumor effect of EcN (Tum-5-p53) was significantly better than that of EcN (Tum-5) and EcN (p53) alone. Because of the powerful capacity of gene editing and packaging, EcN will play an important role in gene therapy.
3.3. Immunotherapy
Cancer immunotherapy aims to activate and harness the body’s immune system to attack malignant cells. However, more than 50% of patients failed to show a durable response to immunotherapy
[30] but have immune-related adverse effects
[31]. There is an urgent need for a targeted, localized, and sustained delivery of immunotherapeutic agents. Programmable EcN provides a convenient approach to meet the demand for immunotherapy. Gurbatri et al.
[32] demonstrated PD-L1 and CTLA-4 antagonists expressed in EcN, enabling continuous and intratumoral checkpoint inhibitor production to induce a durable therapeutic response by a single injection. Due to its local delivery, the risk of systemic toxicities was greatly reduced. EcN has also been studied for its potential utility in vaccine development. Leventhal
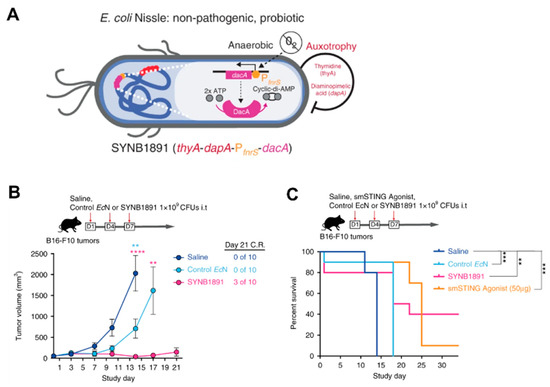
[33] designed an engineering EcN strain named SYNB1891 that expresses the STING-agonist cyclic di-AMP (CDA) to activate antigen-presenting cells (APCs) in the tumor. As shown in
Figure 3, in B16F10 tumor-bearing mice with three i.t. injections over a week, SYNB1891 treatment resulted in complete tumor rejection compared to EcN alone. SYNB1891 treatment also exhibited greater long-term efficacy (40% survival) compared to treatment with the smSTING agonist (10% survival). Furthermore, the cured mice remained tumor-free after re-challenge for at least 60 days. It indicated that SYNB1891 treatment triggered efficacious antitumor immunity and immunological memory.
Figure 3. (A) Schematic illustration of the engineering EcN strain named SYNB1891; (B) Tumor inhibition and (C) survival rates of SYNB1891 treatment in B16F10 tumor-bearing mice. Reproduced with permission from Jose M. Lora, Nature Communications; published by Springer Nature, 2020. ** p = 0.0058 (blue stars), **** p < 0.0001 (pink stars), ** p = 0.0078 (pink stars), ** p = 0.006 (black stars), *** p = 0.0004–0.0006 (black stars).
Localized administration of tumor-targeted EcN provides a distinctive option for tumor immunotherapy, inducing local immune responses to suppress tumor growth while avoiding systemic toxicity. Although the studies of live bacteria in tumor immunotherapy are still in their infancy, it opens up new opportunities and provides guidance for the development of future approaches to improve cancer immunotherapy.
 +1 point
+1 point