3. Discussion
The success of NHP AIDS models has led to increased demand in NHPs in recent years
[18], but appropriate selection of a pre-clinical SIV/SHIV NHP model is critical when evaluating HIV interventions. In these studies, we report less variability in viral control during cART in SHIV-1157ipd3N4 versus SIVΔB670-infected RMs. Specifically, 9/19 SIV-infected animals did not exhibit sustained virologic control during cART compared to SHIV-infected animals which maintained low/undetectable levels of viral replication within a few weeks of starting cART. During treated human HIV infection viral suppression is regularly achieved within 12–24 weeks
[19] and is highly consistent with the responses in SHIV-infected animals reported here. Historically, control of SIVΔB670 replication by antiviral drugs has been highly variable, with some animals unable to control viral replication despite 9 months of treatment
[9]. While we cannot rule out that longer treatment could result in undetectable levels of plasma viremia, our results indicate that greater restoration of mucosal T-cell functions occurs in animals that exhibit more immediate control of viremia after initiating cART. These studies were limited to evaluation of viral burden in the blood. Studies in NHP have shown a relationship between plasma and tissue levels of virus replication
[20][21] and decreased penetrance of antiretrovirals to mucosal sites
[22], so it is likely that animals with high plasma viremia also had greater viral burden at mucosal sites. Future studies are needed to evaluate the viral burden in mucosal compartments to determine the relationship between local viral replication and mucosal CD4 T-cells more conclusively. Antiretroviral drugs designed to inhibit HIV replication can be less efficient in suppressing SIV viral replication and can often take months to achieve durable viral suppression and our findings are consistent with previous reports
[23][24]. Although the classes of cART regimens were similar, studies in humans have revealed higher interpatient pharmacokinetic variability with RAL, that was used in ourSIV infected RMs, and less variability with DTG, that was given to our SHIV infected RMs
[25]. This may explain the increased variability in viral control we see in the SIV-infected RMs. In addition, all three drugs given to the SHIV-infected animals were injectable, whereas one drug (RAL) was delivered orally in the SIV-infected animals; this may have influenced relative cART “adherence” and effectiveness of this regimen in the SIV-infected animals. Alternatively, if the SIV-infected animals were given the same ART regimen as the SHIV-infected animals (e.g., TDF, FTC, DTG) greater viral suppression may have occurred. This specific drug regimen was given to 3 SIVmac239X infected rhesus macaques 2 weeks after infection, yet stable viral suppression was also not achieved until 12–20 weeks following cART
[26]. These data suggest that even if the SIVΔB670-infected RMs had been given a cART regimen of TDF, FTC, DTG, viral suppression may have been incomplete or taken longer than during SHIV infection, thus allowing a greater period to disrupt gut immunostasis.
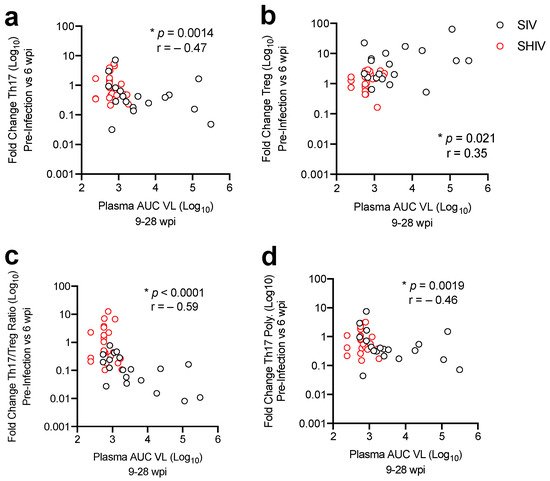
Sustained virologic control during cART, primarily in SHIV-1157ipd3N4-infected RMs, corresponded with less dysregulation of the Th17/Treg ratio during acute infection. These results highlight differences in viral suppression during cART responsiveness in NHP models during cART with 100% of SHIV-1157ipd3N4 and only 53% of SIVΔB670-infected RMs reaching virologic control on cART. Furthermore, better virologic control in response to cART corresponded to less mucosal immune dysfunction, including higher Th17/Treg ratios and lower gut immune activation in SHIV than in SIV-infected animals. These results demonstrate that the SIV and SHIV NHP AIDS models recapitulate aspects of mucosal immune dysfunction found during HIV infection, with signs of increased mucosal pathogenesis in SIVΔB670-infected RMs. Additionally, we found that increased pathogenesis during acute infection leads to greater disruption of mucosal CD4 T-cells and corresponds to poorer virologic control during cART. However, several differences in the experimental design, including virus strain, inoculation dose, time of cART initiation and drug regimen, etc. can influence the mucosal environment, and therefore, differences between the SHIV and SIV models that we report here. Thus, the level of viral pathogenesis not only in the periphery, but also at mucosal sites, needs to be considered when determining whether to use SHIV or SIV NHP models in HIV vaccine studies.
Early cART treatment during HIV infection has been shown to preserve CD4 and Th17 cells in the gut, but has been less successful in reversing persistent gut inflammation, reducing chronic immune activation, or eliminating viral reservoirs
[27][28]. During treated human HIV infection, early cART initiation was found to restore Th17 cell numbers, but not Th17 polyfunctionality and demonstrates how early stages of the infection can have detrimental long-term influences on mucosal T-cell compartments
[29]. Consistent with these findings, we found that mucosal CD4 and Th17 restoration during cART was similar in both infection models, restoration of Th17 polyfunctionality was greater in SHIV infected RMs. Therefore, the SHIV model reported here may better recapitulate aspects of gut mucosal dysfunction observed in cART-treated people living with HIV. This is likely due to the lower viral replication and more efficient suppression of viral replication on cART in the SHIV model. Although cART improves some aspects of HIV infection and disease progression, the findings reported here, including chronic immune activation and dysregulation of Th17 and Treg cells, suggest that strategies to improve gut mucosal homeostasis may increase the effectiveness of immunotherapies aimed to treat or cure HIV infection. Furthermore, our results provide strong evidence that both the SIV and SHIV macaque models of AIDS recapitulate declines in mucosal T-cell function and immune activation, but that the SHIV infection model may more closely mimic this relationship in treated people living with HIV.
Structural barriers and gaps in the HIV care continuum have led to many people living with untreated HIV. As of 2020, 27% of people with a known HIV status did not have access to cART and of those receiving treatment 44% were not virally suppressed
[30]. Failure to achieve long-term HIV viral suppression during cART, due to noncompliance, drug resistance, or other factors, is associated with increased risk of AIDS- and non-AIDS associated co-morbidities
[31] and could ultimately influence the efficacy of HIV intervention strategies. Spontaneous viral remission occurs in untreated individuals
[32] or post-treatment controllers after analytic treatment interruption
[33] and can lead to overestimations in the effectiveness of therapies. Thus, the SIV model, which exhibited greater immune dysregulation consistent with suboptimal cART regimens, may better recapitulate individuals with less accessibility or poorer adherence to cART. Variability of virologic control during cART in pre-clinical NHP AIDS models may impact the relative evaluation of candidate therapeutic interventions for HIV cure in humans and selection of an appropriate NHP model that represents the target population for a given therapeutic is critical. The SHIV macaque model may be better for identifying initial vaccine candidates, but it may be necessary, when possible, to switch to a SIV model, that has greater variability in cART viral control and gut immune dysfunction, to model aspects of real-world scenarios of cART nonadherence or unsuppressed HIV replication.
 +1 point
+1 point