1000/1000
Hot
Most Recent
 +1 point
+1 point
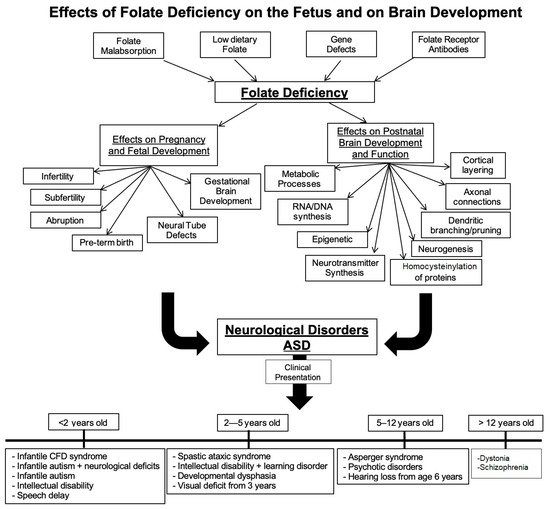
Folate deficiency and folate receptor autoimmune disorder are major contributors to infertility, pregnancy related complications and abnormal fetal development including structural and functional abnormalities of the brain. Food fortification and prenatal folic acid supplementation has reduced the incidence of neural tube defect (NTD) pregnancies but is unlikely to prevent pregnancy-related complications in the presence of folate receptor autoantibodies (FRAb). In pregnancy, these autoantibodies can block folate transport to the fetus and in young children, folate transport to the brain. These antibodies are prevalent in neural tube defect pregnancies and in developmental disorders such as cerebral folate deficiency (CFD) syndrome and autism spectrum disorder (ASD). In the latter conditions, folinic acid treatment has shown clinical improvement in some of the core ASD deficits. Early testing for folate receptor autoantibodies and intervention is likely to result in a positive outcome
| Protein | Gene | Chromosome | GPI Anchor? | Localization | Cofactors? | Refs. |
|---|---|---|---|---|---|---|
| FRα | FOLR1 | 11q13.3 | Yes | Liver, kidney, uterus, placenta, choroid plexus, retinal pigment epithelium | LRP2 | [33][35][36][37][38] |
| FRβ | FOLR2 | 11q13.4 | Yes | Placenta, spleen, bone marrow, thymus, macrophages | NA | [33][35][36][37][38][39] |
| FRγ | FOLR3 | 11q13.4 | NA | Secretory granules of neutrophil granulocytes | NA | [33][35][36][37][38][39] |
| FRδ | FOLR4 | 11q14 | Yes | Oocytes | NA | [33][35][36][37][38][39] |
| RFC | SLC19A1 | 21q22.3 | No | Liver, kidney, placenta, choroid plexus, intestinal tract | Vitamin D, thiamine pyrophosphate | [34][40] |
| PCFT | SLC46A1 | 17q11.2 | No | Liver, kidney, choroid plexus, placenta, intestinal epithelium, human tumors | Proton gradient | [34][40] |





