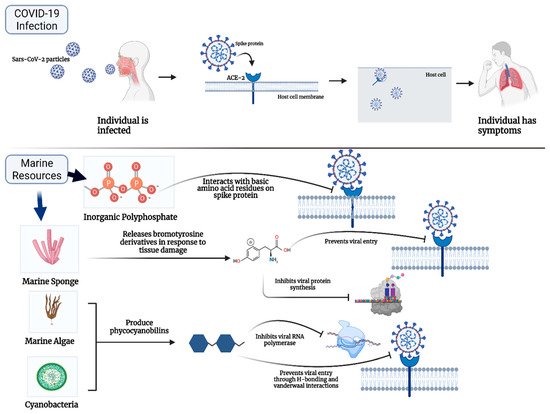
2. Marine Natural Polymer: Inorganic Polyphosphate for COVID-19 Treatment
Inorganic polyphosphate (polyP) is a compound ubiquitously expressed in every cell, including marine organisms, like the cyanobacterium synechcoccus
[20][21][22][23][24]. PolyP is found abundantly in marine bacteria, sponges as well as human blood platelets
[20][21][22][23][24][25]. PolyP, which is released from platelets, interacts with the protease coagulation factor VII and plays an important role in the mediation of blood clots
[20][21][22][23][24][25]. It has been shown that COVID-19 patients have deficient platelet counts and as a result, have reduced polyP in addition to chemical immune mediators such as cytokines and chemokines
[20][21]. Since polyP is abundantly present in platelets, a reduction or deficiency in platelets causes significant reduction in polyP, which can lead to problems with coagulation in Sars-CoV-2 patients
[20][21][22]. Asymptomatic Sars-CoV-2 patients do not have a severe platelet deficiency and thus, it has been proposed that polyP serves a protective role in these patients. PolyP has been shown to bind the RBD of the spike protein on Sars-Cov-2 particles through its basic residues and prevent the binding of the spike protein to host ACE-2 receptors
[20][21][22][23][24] (Refer to
Table 1).
Table 1. Summary of marine compounds for potential Sars-Cov-2 treatment.
| Marine Compound |
Source |
Mechanism of Action |
Inorganic polyphosphate (polyP)
[21][22][24][25] |
Marine sponges, bacteria (ex. Cyanobacterium synepchcoccus) |
- -
-
Binds RBD of spike protein and prevents binding to ACE-2
- -
-
Stimulates innate immune system through upregulation of mucosal proteins
- -
-
Synergistic effects with 4.5 μM dexamethasone or 0.08 g quercetin
- -
-
PolyP120 downregulates ACE-2 by inducing its degradation
- -
-
PolyP120 inhibits Nf-kB pathway and reduces cytokine storm
|
Lambda-carrageenan
[43] |
Marine algae |
- -
-
Reduces expression of viral proteins by suppressing viral replication
|
Terphenyllin
Tirandamycin A
[29] |
Scleractinia associated organisms |
- -
-
Form hydrogen bonds and dock with Mpro
|
Phlorotannins (17 molecules)
[30] |
Sargassum spinuligerum brown algea |
- -
-
Inhibit Sars-Cov-2 Mpro through hydrogen bonding and hydrophobic interactions
|
Five Marine compounds (C19H40O3, C16H30O2, C22H32O4, C21H26O3, C31H30Br6N4O11)
[31] |
Aplysindae Sponge, soft coral Pterogorgia citrina
Petrosia strongylophora sp. |
- -
-
Interact with Mpro through hydrogen and hydropobic interactions
|
Phycocyanobilins (PCB)
[32][33] |
Cyanobacteria, algae rhodophytes |
- -
-
Inhibits Mpro and RNA dependant RNA polymerase
- -
-
Interact with RBD of spike protein through Vander Waal interactions and hydrogen bonding
|
Sulfated Polysaccharides
[34][35][44][45] |
Cyanobacteria, brown algae (Saccharina japonica) |
- -
-
Binds spike protein and prevents viral entry into cells
- -
-
Plays an important role in shielding against the virus due to their anionic properties and molecular weight
|
Bromotyrosines
[40][41][46][47] |
Marine sponges |
- -
-
Inhibits protein synthesis, replication, and proliferation of HIV-1
- -
-
Binds spike protein and inhibits viral entry
|
In particular, one study synthesized a model depicting the proposed mechanism by which polyP interacts with the spike protein
[24]. Approximately 15 phosphate units of polyP are thought to interact with the basic residues, Arg, Lys and His on the spike protein
[20][24]. Furthermore, the study found that the soluble polyP significantly inhibited the interaction of the spike protein and Ace-2 at concentrations ranging from 1 μg/mL to 100 μg/mL
[24]. This inhibition was found to be 70% effective, suggesting that polyP has protective anti-viral effects
[24]. Notably, during 24 h of incubation, polyP ranging up to 100 μg/mL had no toxic effect on cells.
Further, polyP is typically hydrolyzed by alkaline phosphatase (ALP) which releases free energy and results in the formation of ADP which becomes phosphorylated to form ATP
[21][22]. It has been found that polyP is able to stimulate the innate antiviral immune response by inducing mucin gene expression through the increased generation of ATP
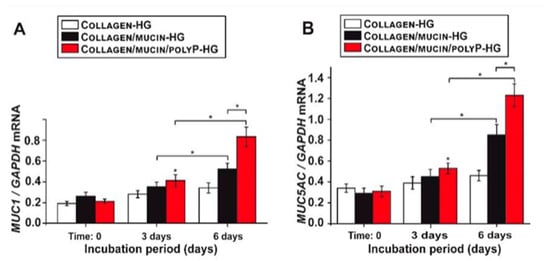
[21][22]. As seen by
Figure 2, compared to the controls (black and white), when polyP was added, mRNA levels of MUC1, a main mucin type, significantly increased over 6 days compared to controls
[21]. These findings suggest that in addition to preventing Sars-Cov-2 attachment to host cells, polyP boosts the innate immune system and mucosal defense against the virus. This is significant as one’s status of innate immunity drastically contributes to the manifestation of COVID-19 in patients.
Figure 2. Expression of MUC1 and MUC5AC genes in A549 cells over 6 days. (
A) Relative expression of MUC1 incubated for 3 and 6 days. (
B) Relative expression of MUC5AC incubated for 3 and 6 days
[21].
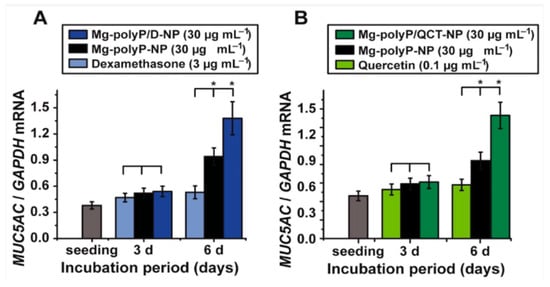
Similarly, another study utilized nanoparticles of polyP in addition to dexamethasone or the metabolite quercetin on A459 epithelial cells
[22]. The study found that polyP increased the expression of a major mucus glycoprotein MUC5AC
[22]. As seen in
Figure 3, polyP combined with 4.5 μM dexamethasone (anti-inflammatory) or 0.08 μg quercetin (antioxidant) significantly increased the expression of MUC5AC than either drug alone. Dexamethasone causes some toxic effects (induction of apoptosis) at concentrations greater than 100 mM, thus the 4.5 μM used in the study did not affect cell viability. Similarly, quercetin is cytotoxic at concentrations greater than 0.3 μg/mL, thus the 0.08 μg used in the study did not impact cell viability
[22]. These synergistic effects demonstrate the potential of combinatory therapy involving both marine resources and non-marine drugs. MUC5AC and MUC1 genes encode for proteins that play an important role in the mucosal barrier and are shown to be elevated in healthy individuals in comparison to Sars-Cov-2 patients
[22]. This is significant as the mucosal barrier is a critical part of defense against Sars-Cov-2 and other pathogens, as it clears the viral particles and dictates the accessibility of the pathogens to the host epithelial cells.
Figure 3. Expression of MUC5AC genes in A549 cells. (
A) Cells were exposed to 3 μg mL
−1 of dexamethasone, 30 μg mL
−1 of polyP and dexamethasone (“Mg-polyP/D-NP”) or 30 μg mL
−1 of polyP (“Mg-polyP-NP”). (
B) Cells were exposed to 0.1 μg mL
−1 of quercetin, 30 μg mL
−1 of polyP and quercetin (“Mg-polyP/QCT-NP”) or 30 μg mL
−1 of polyP (“Mg-polyP-NP”)
[22].
In line with these studies, Ferrucci et al. demonstrated that polyP120 binds to the ACE2 receptors and downregulates it by inducing its degradation
[25]. As seen by Ferrucci et al., immunoblotting demonstrated a dose-dependent decrease in ACE-2 expression as the concentration of polyP120 increased over 24 h
[25]. This is promising as the mode of Sars-Cov-2 entry is through the ACE2 receptor on host cells. Furthermore, the study demonstrated that polyP120 impairs the synthesis of viral proteins required for the replication of Sars-Cov-2 by impairing viral transcription and replication
[25]. The study also found that the inorganic polyphosphate inhibits the Nf-kB pathway and thus, reduces the cytokine storm typically associated with COVID-19 infection
[25]. This is valuable as systemic infection and significant inflammatory immune response can be detrimental.
3. Promising Compounds from Marine Algae, Bacteria, Sponges, and Fish for COVID-19 Treatment
Marine algae are known to be a source of numerous bioactive substances such as vitamin E, B12, phycocyanin, lutein and polysaccharides
[48][49][50]. Specifically, lambda-carrageenan is a polysaccharide purified from marine red algae (Refer to
Table 1) and has anti-viral, anti-bacterial, anti-cancerous and anti-coagulant functions
[48][49][50]. It has been shown to effectively inhibit both influenza virus and Sars-Cov-2
[43]. A study done by Jang et al. demonstrated that the marine polysaccharide was able to reduce the expression of viral proteins and suppress viral replication dose-dependently
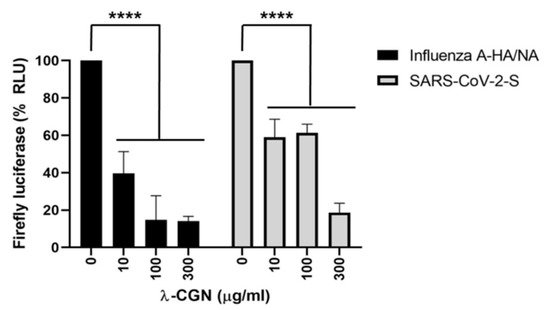
[43]. Depicted in
Figure 4, as the dose of lambda-carrageenan increased from 0 to 300 μg/mL, the presence of spike viral proteins on Sars-CoV-2 and influenza A viral proteins significantly decreased
[29]. Inhibition of Influenza virus and Sars-Cov-2 depicted EC50 values 0.3–1.4 μg/mL and 0.9 ± 1.1 μg/mL, respectively. Favorably, no host cell toxicity was observed at concentrations up to 300 μg/mL
[29]. The study also found that mice challenged with Sars-Cov-2 virus and later given Lambda-carrageenan had a 60% survival rate, as the polysaccharide inhibited viral entry and viral replication
[43]. These findings reveal the anti-viral properties of lambda-carrageenan and make it a promising marine resource for COVID-19 treatment.
Figure 4. Inhibition of Influenza A and Sars-CoV-2 by lambda-carrageenan expressed as firefly luciferase. Influenza A viral proteins (black bars) over increasing concentrations of lambda-carrageenan. Sars-Cov-2 spike proteins (grey bars) over increasing concentration of lambda-carrageenan
[43].
Although these findings are promising, it is important to mention potential adverse effects of lambda-carrageenan. Previous studies have reported the oligosaccharides derived from the carrageenan family (kappa- and lambda-carrageenan) can impair blood vessel development by inhibiting the growth of new blood vessels
[48][51]. It was also found that at 200 μg/mL they could inhibit migration, proliferation as well as tube formation of human umbilical vein endothelial cells
[48][51]. These results demonstrate potential toxic effects to humans; however, more in vitro and in vivo toxicology studies are needed. It is important these studies be taken into consideration for further development of lambda-carrageenan against Sars-Cov-2.
The pharmacological potential of sea organisms is further prevalent in scleractinia associated organisms such as bacteria and fungi
[13][14][15][26][27][28][29]. These organisms are known to produce a variety of metabolites making them implicated in inflammation and viral infection
[13][14][15][26][27][28][29]. In a study done by Zahran et al., scleractinia related metabolites were analyzed, and molecular docking was performed to determine potential Sars-CoV-2 anti-viral effects
[29]. It was found that two specific microbial metabolites (Terphenyllin and Tirandamycin A) form hydrogen bonds and dock with high affinity to the main protease (M
pro)
[29]. These marine metabolites are thought to be promising leads for inhibition of the main protease, which plays an important role in the life cycle of the virus. Similarly, Gentile et al. identified seventeen potential M
pro inhibitors from the class phlorotannins isolated from
Sargassum spinuligerum brown algea
[30]. Docking energies ranged from −14.6 to −10.7 kcal/mol and the compounds interacted with M
pro through extensive hydrogen bonding as well as hydrophobic interactions. In addition, the Sars-Cov-2 RNA polymerase along with nsp7/8 are required for the RNA replication and viral protein synthesis
[30]. Remdesivir is a known inhibitor of the RNA dependent-RNA polymerase, and three marine Scleractinia metabolites were found to bind the polymerase in the same position as Remdesivir
[6][7][8][30]. This finding suggests that these marine metabolites are promising leads for the inhibition of viral replication and thus, treatment of COVID-19.
Moreover, a study done by Khan et al. performed molecular docking analysis on M
pro and found that several marine compounds demonstrated promising binding interactions
[31]. Five marine compounds, isolated from sea sponges of family Aplysinidae and
Petrosia stronglyophora sp. and the soft coral
Pterogorgia citrina were found to interact with M
pro through hydrogen and hydrophobic interactions
[31]. Analysis of ADME properties depicted them to have potential Sars-Cov-2 therapeutic application
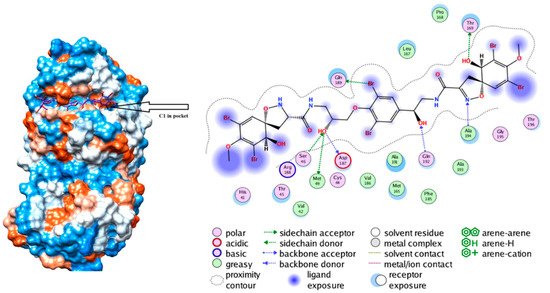
[31]. One marine compound (C1, from the family Aplysinidae) was found to fit the M
pro pocket the best, having affinity for all regions of M
pro with much higher hydrogen and hydrophobic interactions
[31]. As seen in
Figure 5, C1 residues Ser46, Met49, Asp186, Gln192, Ala194, Thr169 as well as Gln189 interacted with M
pro through H-bonding
[31]. This finding gives insight into the spatial location that the compounds have in the binding pocket where there are also hydrophobic and electrostatic interactions present.
Figure 5. Depicts the interaction of C1 compound with Sars-Cov-2 M
pro after molecular dynamic stimulation. C1 hydrogen bonds with Ser46, Met49, Asp186, Gln192, Ala194, Thr169 and Gln189. Reproduced with permission from Khan, M.T. et al., Marine natural com-pounds as potents inhibitors against the main protease of SARS-CoV-2-a molecular dynamic study; published by Taylor & Francis, 2020,
[31].
Additionally, phycocyanobilins (PCBs) are pigment compounds present in some types of cyanobacteria as well as the algae rhodophytes
[32][33][44]. They have been shown to have antioxidant as well as anti-viral properties making them promising leads for COVID-19 therapy. One study done by Pendyala and Patras demonstrated that the marine PCB among others, are potent inhibitors of M
pro and RNA dependant RNA polymerase (RdRp) of Sars-Cov-2
[32]. Through in-silico screening, it was depicted that the PCBs had a higher binding affinity to RdRp than the current drug Remdesivir which signifies the potential these compounds have for anti-Sars-Cov-2 effects
[32]. Similarly, Petit et al. performed an in-silico study and found that PCB among other phycobilin compounds expressed by
Arthrospira had promising anti-viral properties against Sars-Cov-2
[33]. The study found that PCB interacted with the RBD of the virus’ spike protein through Vander Waal interactions as well as hydrogen bonding. PCB was found to have a competitive binding energy of −7.2 kcal/mol suggesting it to be a potential anti-viral compound
[33]. Promisingly, the study reported phycobilin compounds from
Arthrospira to have minimal to no cytotoxicity to cells and was shown to be effective at low doses (1–10 μg/mL). PCBs were reported to have low mutagenicity, carcinogenicity and reprotoxicity
[33]. These findings demonstrate that PCBs have substantial anti-viral effects and may serve as promising agents against Sars-Cov-2.
The resources marine organisms provide are never ending. Cyanobacteria harbor numerous metabolites such as sulfated polysaccharides which are known to have anti-viral properties
[34]. Sulfated polysaccharides have anti-viral activity against HSV, hepatitis B virus as well as retroviruses
[34][35][45]. They have been shown to play an important role in shielding against the virus due to their anionic features as well as molecular weight which together are able to have anti-viral effects
[52]. Due to these anti-viral abilities, it is proposed that the polysaccharides harbor large potential against Sars-Cov-2
[34][35][45][46]. In fact, a study done by Kwon et al. demonstrated that a specific type of sulfated polysaccharide, Fucoidan, from
Saccharina japonica, had anti-viral activity against Sars-Cov-2
[35]. The study showed that the marine compound was more potent than Remdesivir, suggesting that it is a promising therapeutic agent against COVID-19
[35]. Similarly, a study done by Song et al. demonstrated that fucoidan from brown algae, cucumber sulfated polysaccharide as well as carrageenan from red algae all displayed anti-viral properties at concentrations ranging from 3.9 to 500 μg/mL
[45]. It was found that the cucumber sulfated polysaccharide had the strongest inhibitory effects due to its ability to bind the spike protein and inhibit viral entry into cells
[45]. Favorably, at concentrations up to 500 μg/mL, no cytotoxicity was observed as depicted by no significant changes in cell viability
[45]. These findings depict the potential that sulfated polysaccharides have for effective treatment of Sars-Cov-2.
As recently proposed by Nguyen and colleagues, “marine sponges have the capacity to filter large volumes of ‘virus-laden’ seawater through their bodies and host dense communities of microbial symbionts, which are likely accessible to viral infection”
[53]. These organisms with typical filter-feeding lifestyles are constantly in contact with the surrounding environment where one milliliter of water can contain up to 10 million viruses that move through their body per day
[53]. Marine
Halichondria panicea demosponge has been shown to rapidly filter out viruses at high rates (176 mL h
−1 g tissue dry wt
−1)
[54]. Thus, it is not surprising that investigations on viral ecogenomics across the four sponge classes is in trend
[55]. Despite predation
[56], sponges develop highly specialized chemical defense mechanisms based on the biosynthesis of effective antiviral biomacromolecules
[57][58]. Such antiviral substances of poriferan origin such as nucleoside Ara-A (vidarabine) isolated from sponge
Tethya crypta [59], or spongouridine and spongothymidine
[60], have been recognized recently. Sponge extracts which have been shown to be active against human adenovirus (HAdV) have also been reported
[61]. After 2019, special attention has been payed to poriferan inhibitors of SARS-CoV-2 key target proteins (i.e., Ilimaquinone)
[36], or Remdesivir
[37].
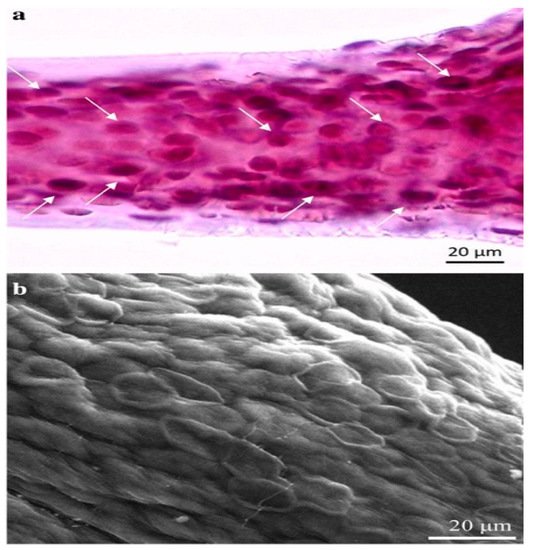
Marine demosponges, which belong to the Verongiida order, are recognized producers of bromotyrosines
[38][39][40]. They possess anti-viral, anti-bacterial and anti-parasitic properties
[40][41][42][46]. These brominated compounds in the aplysinidae family of verongiids are produced within specialized cells known as spherulocytes (
Figure 6)
[47]. These cells are sensitive to changes in the environment and can release bromotyrosine in response to environmental stimulus
[47].
Figure 6. Image depicts the specialized spherulocyte cells in the chitinous skeletal fibers of verongiid sponges. (
a) Light microscopy image showing the distribution of these specialized spherulocyte cells (arrows). (
b) Scanning lectron microscopy represents the morphology of these cells. Reproduced with permission from Muzychka, L. et al., Marine biomimetics: Bromotyrosines loaded chitinous skeleton as source of antibacterial agents; published by Springer Nature, 2021,
[47].
Recently, Muzychka and co-workers isolated the bromotyrosine derivative 3,5-dibromoquinolacetic acid using a novel biomimetic water-based method and found it to have anti-microbial properties against selected clinical stains of
Staphylococcus aureus,
Enterococcus faecalis and
Propionibacterium acnes [47]. The study also showed that Aeroplysinin-1 and 2 also had anti-microbial properties
[47]. Similarly, another study was able to isolate the bromotyrosine Aeroplysinin-1 and found that it was able to decrease the viability of neuroblastoma cell lines and inhibit the growth of drug resistant bacteria
[41]. Similarly, bromotyrosines have been shown to have potential anti-tumor and anti-metastatic affects displaying cytotoxic properties
[40]. Dreschel et al. investigated the potential cytotoxicity and therapeutic window of two bromotyrosine derivatives, Aerothionin and Homoaerothinin
[40]. The study found that after treating mouse endothelial cells with 25 to 50 μM Aerothionin or Homoaerothinin, cell viability was significantly reduced. However, the bromotyrosine derivatives had efficacious anti-cancerous effects at 10 μM of Aerothionin or Homoaerothinin, with minimal cytotoxic effects
[40].
Moreover, bromotyrosines have also been found to inhibit HIV-1 infection through inhibiting protein synthesis, replication and proliferation of the virus as well as preventing its entry into host cells (refer to
Table 1)
[42]. This is promising as bromotyrosines may be useful for treating COVID-19. In a recent press release, Dr. Ehrlich emphasized that his research group was able isolate reasonable amounts of bromotyrosines which can be used to further investigate their potential against the Sars-Cov-2 virus
[46]. Thus, given bromotyrosines anti-viral and anti-pathogenic effects, future clinical trials investigating it further are valuable for potential COVID-19 treatment.
4. Promising Advantages and Limitations of Marine Resources
The resources marine organisms harbor are limitless and consistently prove efficacious at combatting viruses, bacteria, cancers and other pathogens. Their unique chemical structures and diversity introduce novel mechanisms of action, making them especially valuable against drug-resistant pathogens. Some marine compounds that do share similar mechanisms of action with other known approved drugs have shown to be more potent. As discussed above, PCBs and sulfated polysaccharides have shown to bind and inhibit RdRp with higher affinity than current standard therapy Remdesivir
[27][30]. Furthermore, each marine compound serves not one single role, rather multiple roles proving valuable for different applications. For example, the PCBs are not only able to inhibit RdRp, but also interact with the RBD of the viral spike proteins, making them even more advantageous over Remdesivir
[33]. In addition, majority of the compounds such as sulfated polysaccharides do not only possess anti-viral properties, but also anti-coagulant, anti-inflammatory, anti-oxidant and anti-bacterial
[62]. These multi-faceted properties of marine compounds make them very efficacious agents against Sars-Cov-2. This is advantageous over synthetic compounds which typically possess one single valuable property and are often administered in combinatory therapies, which increases the possibility of drug-drug interactions. In addition, due to the abundance and diversity of marine resources, they are highly cost-effective. This makes them valuable, as the current standard treatment Remdesivir costs approximately $2600 for a 5-day course treatment
[63]. Furthermore, at effective concentrations of polyP (<100 μg/mL), lambda carrageenan (<300 μg/mL), PCBs (<10 μg/mL), sulfated polysaccharides (<500 μg/mL) and bromotyrosines (10 μM), no toxic effects on cells were observed
[21][33][40][43][45].
However, the process of marine drug development is faced with many challenges. Firstly, although the sea harbors countless organisms, accessibility to majority of these resources are limited
[64]. Although plentiful compounds are accessible close to shore, there remain other regions of the ocean that likely possess unknown organisms and thus, new therapies
[64]. Furthermore, to continue the development of promising compounds through pre-clinical and clinical trials, there must be a continuous supply of the compounds. This presents a challenge as large-scale production may harm the marine ecosystem
[64][65]. Fortunately, rapid technological advancements in synthetic chemistry and biotechnology provide a potential solution to this problem
[64]. In addition, many potential anti-viral metabolites have only been tested in vitro or visualized through molecular docking assays. More in vivo studies are needed to further investigate potential adverse effects and drug delivery requirements. Despite the challenges faced, it is clear that marine organisms serve as a promising avenue for future pharmacological intervention.
 +1 point
+1 point