1. Introduction
Eucalyptus (family Myrtaceae) is a large genus of fast-growing evergreen trees and ornamental shrubs with more than 900 species, native to Australia, Indonesia, India, Portugal, South Africa, Brazil, Chile, and France
[1]. The economic importance and commercial value of
Eucalyptus have long been considered in many parts of the world due to its hard timber, resistance to biotic stresses, rapid growth, and high profitability
[2][3][4].
Furthermore, increasing global demand for timber, pulp, and paper products has significantly changed the plantation culture of
Eucalyptus species all over the world
[5]. Besides having extensive use of
Eucalyptus in agroforestry systems, the essential oils extracted from this tree have been widely used in modern ethnopharmaceutical studies due to its various biological activities and medicinal properties
[6]. To date, natural and cultivated
Eucalyptus forests have been considered as the initial genetic resources for agroforestry plans and tree breeding programs
[7]. Despite the abundance of natural
Eucalyptus forests, cultivation of
Eucalyptus by seed is the most traditional way of propagation with varying degrees of competitive success and establishment. In the latter stage, vegetative propagations methods have also been applied for
Eucalyptus improvement programs
[8][9]. Vegetative progeny methods are broadly being used for the asexual propagation of forest trees. The primary vegetative propagation techniques are grafting, layering, root cuttings, and rooting of the shoot. Generally, the effectiveness of the methods depends on the physiological structure, shape, performance, and age of the donor tree
[10]. Over recent decades, conventional breeding tools which have been implemented in forestry improvement programs have become the main strategy to multiply forest trees
[11]. However, the bottlenecks of conventional breeding techniques opened a new window to the clonal propagation of
Eucalyptus.
Clonal propagation is often considered as one of the most critical
in vitro platforms to increase the multiplication rate of
Eucalyptus sp. in forestry breeding programs and biotechnology research
[12]. Micropropagation, which is mainly achieved via somatic embryogenesis and organogenesis, is the true-to-type, virus-free, and rapid propagation technique
[13]. The
in vitro culture of
Eucalyptus depends on several factors including external aspects (sugar concentrations, pH, media composition, plant growth regulators (PGRs), luminosity, and temperature) and internal features (physiological conditions, age, and genotype)
[13]. Despite the advancement in the micropropagation method, the obstacles in callus induction, regeneration, root induction, and acclimatisation are still the major challenges for clonal propagation of
Eucalyptus. On the other hand, hyperhydricity, rapid browning, and poor explant response, which cause poor growth of explants and even failure of tissue culture procedure, are some of the other major factors affecting
in vitro propagation of
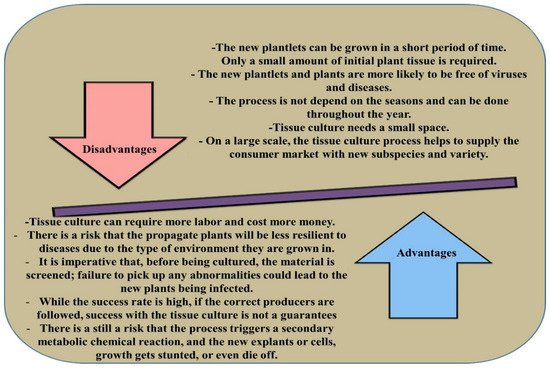
Eucalyptus [14]. The advantages and disadvantages of micropropagation methods in plants are presented in
Figure 1 [15].
Figure 1. The advantages and disadvantages of micropropagation in plants.
To date, several micropropagation techniques and in vitro protocols have been developed for tissue culture of Eucalyptus. Regardless of the advances, micropropagation of Eucalyptus has been met with limited success (failures) and applicability. This is due to the lack of studies focused on the mechanisms, strategies, and interactions of internal and external factors under in vitro conditions. Keeping this in view, this current review aims to clarify the rationale mechanisms behind the observed phenomenon in Eucalyptus tissue culture. Furthermore, this is an extensive review of some intriguing aspects of internal mechanisms of Eucalyptus in response to external (Physico-chemical) factors under in vitro conditions.
2. Organogenesis
Organogenesis is the vegetative propagation by which plant organs such as flower buds, shoots, and roots are produced from the cells and tissues (the unusual points of origin) under
in vitro conditions. The type of organogenesis (direct and indirect) depends on the presence of relative concentrations of hormones and explants in the culture medium
[16]. Indirect organogenesis is the formation and development of organs from an amorphous tissue, callus, or cell suspension. Additionally, this propagation method is a reliable technique, which has been applied for the production of genetically modified plants from calli
[17]. On the other hand, the development of new organs from the explants (axillary buds), which forms new shoots and buds, is known as direct organogenesis
[18].
In
Eucalyptus, the composition of different factors such as carbon, various vitamins, amino acids, gelling agent, macro-and micronutrients, and other additives has affected the efficacy of both types of organogenesis
[19]. For instance, during indirect organogenesis of
Eucalyptus camaldulensis hybrids, the combination of naphthyl acetic acid (NAA) and 6-benzylaminopurine (BAP) showed a better efficacy as compared to the application of zeatin, kinetin, casein hydrolysate, and 2,4-Dichlorophenoxyacetic acid (2,4-D)
[20]. Additionally, callus induction was observed in
E. camaldulensis hybrids cultivated on Murashige and Skoog Basal (MS) media added with 1 mg L
−1 NAA, and the highest rate of somatic embryogenesis was achieved on MS basal medium fortified with 0.1 mg l
−1 NAA and 0.5 mg l
−1 benzyladenine (BA)
[21].
3. Somatic Embryogenesis
Somatic embryogenesis (SE) is a multifactorial, non-sexual, and complex biotechnological tool. This artificial method produces bipolar embryos through physiological, biochemical, and molecular pathways from somatic tissues. During SE, somatic cells of plants become totipotent and alter their pathway of development, resulting in the formation of a complete plant from somatic embryos
[22]. In plants, two arrays of the somatic embryos have been reported including indirect somatic embryogenesis induced from an unorganised callus and direct somatic embryogenesis induced from the pre-embryogenic cells (cells of the nucellus)
[23]. Indirect embryogenesis needs redetermination of differentiated cells process, callus induction, proliferation, and the development of the embryogenically determined phase
[24]. Generally, the effects of suitable PGRs at specific concentrations play an integral role in the re-entry of cells into mitosis and determination of the embryogenic phase
[25]. Nonetheless, during direct somatic embryogenesis, which is a rare type of SE, the cell(s) produces embryos without the formation of an intervening callus. To achieve direct embryogenesis, pre-embryogenic determined cells (PEDC) require favourable conditions and specific type of PGRs to enter mitosis and complete embryogenesis
[26].
Over the decades, numerous somatic embryogenesis protocols for several
Eucalyptus species have been reported
[13][27][28]. However, the occurrence of somaclonal variation, low percentage of embryogenic initiation, and an inability of somatic embryos to reach complete maturation limit somatic embryogenesis adoption for clonal propagation in
Eucalyptus [14]. It has been also reported that several factors, such as hormonal type and level, culture media, and ontogenetic age of tissue interfere with the diverse phases of somatic embryos
[29]. Nonetheless, there is a notorious paucity of studies addressing ultrastructural, histological, and cytological evidence on the diverse features linked to the frequency induction and proliferation of somatic embryos. Despite the morphological resemblances of somatic embryos with zygotic ones at the proliferation and developmental phases, a lack of clear definition has been stated in some other phases of proliferation and developmental steps in
Eucalyptus [30]. For example, histological and morphological investigations on the embryogenic cell of
Eucalyptus globulus,
Eucalyptus grandis, and
Eucalyptus nitens at diverse development phases demonstrated the usual features of other somatic embryogenic structures including small vacuole, prominent nucleus, small volume, and dense cytoplasm
[31].
The success rate of a propagation technique in Eucalyptus depends on both quality (genetic stability and growth rate) and quantity (survival) of the regenerated plants. The combination of the above-mentioned factors associated with the genotypes of Eucalyptus is the most effective aspect in both somatic embryogenesis and organogenesis. In the following sections, the effects of the most important factors on both somatic embryogenesis and organogenesis of Eucalyptus are discussed.
 +1 point
+1 point