2. The LPA3 Receptor: Structure and Function
The human LPA
3 receptor (
https://www.uniprot.org/uniprot/Q9UBY5; Accessed on 12 May 2021) is constituted of 353 amino acids (mouse and rat orthologs, 354 amino acids), and its calculated molecular weight is ≈40 KDa (39,998 Da)
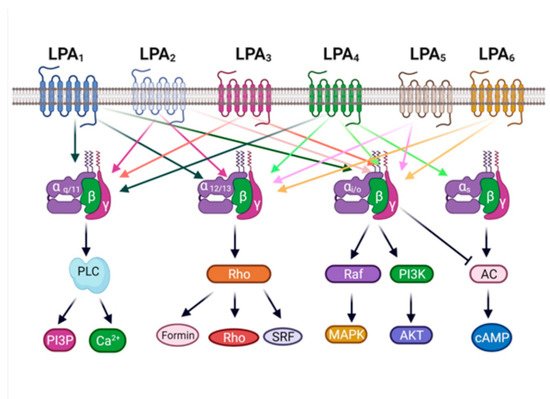
[5][55][56]. As previously indicated, according to the classification systems GRAFS and A-F, this receptor belongs to the A family
[36][37]. LPA
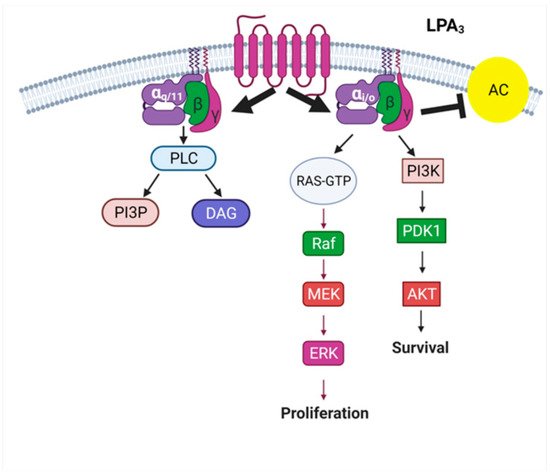
3 is mainly coupled to two G proteins, Gα
q/11 and Gα
i/o; therefore, the G protein-binding motif of this receptor subtype is considered promiscuous. This property allows this receptor to activate different signal pathways, which might explain why it does participate in a large variety of physiological functions and, as previously mentioned, in the pathogenesis of diseases
[5][8][57].
As a member of the GPCR superfamily, the LPA
3 receptor is constituted of seven hydrophobic transmembrane domains (TM), which are joined through three extracellular and three intracellular loops (
Figure 5). It is worth mentioning that transmembrane regions are essential for this receptor, as has been observed for others that also belong to the A family. These regions or domains are frequently conserved
[58].
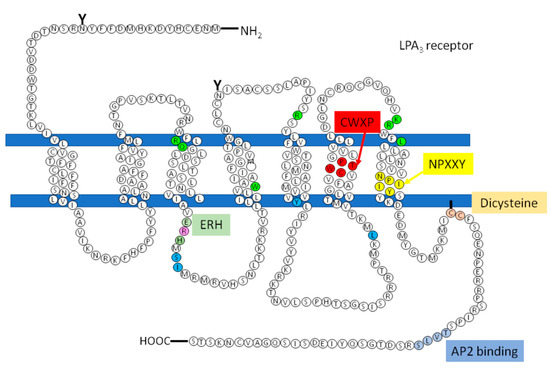
Figure 5. LPA3 receptor structure, domains and sites that regulate this receptor. Image shows the amino acid sequence and the organization of the LPA3 receptor with three extracellular loops, three intracellular loops, the seven transmembrane domains, the extracellular amino terminus (-NH2), and the intracellular carboxyl terminus (-COOH). Colored boxes indicate conserved motifs putatively relevant for activation and regulation of the LPA3 receptor. Putative sites where LPA interacts with LPA3 are shown in green, while proposed places where GPCRs could be recruiting G proteins are marked in blue and purple (R, arginine that is also part of the ERH motif). “Y” indicates a potential glycosylation site, and the line joining one of the cysteines to the membrane is a putative palmitoylation site.
Available information on LPA
3 receptor structure/function is scarce. Therefore, in order to obtain some information, we performed
in silico analyses. This allowed us to identify different domains observed in other GPCRs. Among these are the following: an ERH (Glutamic acid-Arginine-Intrahelical hydrogen bonding residue) domain (analogous to the DRY (Aspartic acid-Arginine-Tyrosine) motif) in the transition between the end of TM3 and the initiation of ICL2, a CWXP domain within TM6, an NPXXY domain near the end of TM7, and a di-cysteine domain within the carboxyl terminus (
Figure 5). Studies on these domains in other receptors have shown that they are important for the activation and regulation of the GPCRs receptors of the A family
[59][60][61][62]. Additionally, an AP2-binding domain is present in the carboxyl terminus
[62][63][64].
It is noteworthy to mention that the mutation of these domains usually reduces or abolishes agonist-activation of GPCRs. Studies employing molecular docking showed that ligand binding at GPCRs produced the packaging of TM3-5-6-7 domains; this event was promoted by destabilization of an ionic interaction
[60][65], initiating a displacement of TM7 toward TM3 and promoting activation involving the tyrosine residue present in the DRY motif, which is associated with the rotation of the cytoplasmic extreme of TM6 and which promotes the activation of these receptors
[60][66][67][68][69].
Additionally, the asparagine residue of the NPXXY motif establishes interactions with other residues, facilitating the movement of TM7 toward TM3
[60][67] and promoting the stability of the activated receptor. Finally, the DRY motif forms a salt bridge with surrounding residues and with TM6; this salt bridge breaks at the moment the ligand binds. The DRY motif creates a new interaction with TM5, stabilizing the receptor in its active conformation, breaking contacts between TM3 and TM6, thus promoting a movement toward the cellular cytoplasm of TM6, which increases the receptor binding to the G
α protein. These events initiate signaling, favor receptor phosphorylation, and later favor association with β-arrestins, all of which are relevant for receptor desensitization
[53][60][67][68][69].
The CWXP domain is a motif found in TM6 which seems to participate in the binding of agonists. Rotation of the tryptophan residue causes movements within the binding pocket, promoting the accommodation of the ligand into the receptor. In contrast, the proline residue induces a bend that serves as a pivot for helical movement during receptor activation
[60][61][67][68][69][70]. Other motifs that appear to participate in the activation of GPCRs include the PIF (GPCR microswitch; Proline-Isoleucine-Phenylalanine) motif that is usually found in TM4 and the NPXXY motif found in TM7, both of which are also related to the activation of Gα
q, Gα
s, Gα
i and β-arrestins
[67][71][72][73][74]. It has been shown that in some receptors (such as the histamine 2 receptor
[74][75], the formyl peptide receptor
[47][68][75], and α- and β-adrenoceptors
[61][76], among others), this domain could be regulating agonist-induced internalization, which affects MAPK pathway activation and intracellular calcium mobilization.
The majority of the motifs that generally regulate the activation of GPCRs, including those in the LPA1 receptor, have also been found in the LPA3 receptor (Figure 5). Only the PIF domain could not be found in the receptor sequence. Therefore, it appears likely that other receptor region(s) could replace the role of PIF in receptor activation.
This illustrates the putative importance of the motifs present in the LPA
3 receptor at the time of its activation when the ligand binds to it; however, we must recall that the intracellular loops and the carboxyl-terminal region play essential roles, particularly in receptor desensitization and internalization. Current ideas suggest key roles in the phosphorylation of specific residues, mediated by GRKs, second messenger-activated, and other protein kinases
[68][77][78].
Other important regions of the LPA
3 structure are the transmembrane domains, which contain residues that take part in ligand binding. It is worth mentioning that the LPA receptors that belong to the lysophospholipid subfamily entertain an ≈81% similarity among themselves
[79][80].
Few studies have reported the participation of these residues during the binding of the ligand in LPA receptors. The residues where LPA has been shown to interact with LPA receptors include arginine 105, glutamine 106, tryptophan 153, arginine 185, lysine 279, and arginine 276 (
Figure 5, residues in green). These sites are conserved in the LPA
1, LPA
2, and LPA
3 receptors, but differences appear to exist between these
[5][55][57][79][80]. In the case of tryptophan 153, when it was mutated to alanine in the LPA
3 receptor, it induced a decrease in the potency and efficacy of LPA; such changes were not observed when the LPA
1 and LPA
2 receptors were similarly mutated. Likewise, when arginine 279 was substituted with alanine, a decrease in the activation of LPA
1 and LPA
2, but not in the LPA
3 receptor, was observed
[79][80].
Another structure important is an amphipathic α-helix, frequently denominated helix 8, that maintains the F (R/K) XX (F/L) XXX (L/F)sequence that is conserved in GPCRs of the A family and has been reported to participate in the maintenance of the receptor on the cell surface promoting GPCR trafficking, and participating in the activation of the G proteins and the receptor’s interaction with the β-arrestins[81][82][83][84].
However, there are receptors of the same family that do not present this sequence that could be involved in the recruitment of the G protein, how is the LPA3, so according to studies carried out by Zhou and coworkers, in which it is proposed that in response to agonist-induced conformational changes, residues in transmembrane domains 3, 5, and 6 interact with and activate G proteins[77]. These residues were found in the structure of the LPA3 receptor as shown in Figure 5 (indicated in cerulean).
The GRKs are a family of protein kinases that appears to play a major role in the phosphorylation of agonist-occupied GPCRs (
Table 1). This family is made up of seven different isoforms that are constituted of a central catalytic domain which is conserved in all GRKs; an amino-terminal area and the carboxyl terminus, both of which differ among these protein kinases, seem to confer them selectivity in their action, and participate in their regulation. These domains constitute the structural basis for their classification into subfamilies; in addition, some GRKs exhibit selective expression in some tissues
[85][86][87][88]. The visual GRKs (GRK1 and GRK7) are mainly expressed in the retina, GRK4 is mainly expressed in the testis, whereas the other GRKs (2, 3, 5, and 6) are ubiquitously expressed; visual GRKs have short prenylation sequences (see reviews in
[85][88] and references therein). The second subfamily, denominated GRK2 and also, for historical reasons, the β-adrenergic receptor kinase (or βARK) subfamily, exhibits a Pleckstrin homology domain that interacts with G protein βγ dimers and phosphatidylinositol 4, 5-bisphosphate. These kinases are cytoplasmic and their interaction with the plasma membrane seems to occur through these domains. The GRK4 subfamily seems to be bound to the plasma membrane through palmitoylation and/or the presence of positively charged lipid-binding elements
[85][86][87][88]. It has been proposed that lipids covalently bound to the carboxyl terminus of these proteins, the Pleckstrin homology domain that associates with phosphoinositides, and the polybasic/hydrophobic regions permit these kinases to be recruited to the membrane and to catalyze GPCR phosphorylation at specific residues
[87][88][89][90][91].
Table 1. GRKs that putatively phosphorylate different sites in GPCRs.
| Subfamilies |
GRKs |
Domains of Interest |
| Visual GRKs |
GRK1 and GRK7 |
Prenylation |
GRK2 or βARK
GRK4 |
GRK2 and GRK3
GRK4, GRK5 and GRK6 |
Pleckstrin homology
Palmitoylation, polybasic hydrophobic domains |
Such specificity in the GPCR phosphorylation pattern appears to be critical to define subsequent signaling (frequently associated with β-arrestin activation), vesicular trafficking, and the receptor’s fate (rapid or slow recycling to the plasma membrane, or degradation). This has been named the “GPCR phosphorylation barcode,” and numerous research groups are actively working to understand (i.e., to break) this code, which currently is only partially understood
[46][50][92][93][94][95][96]. Obviously, initial steps include knowing that the GPCR of interest is actually phosphorylated, the conditions under which that takes place, and the definition of the specific sites affected by such covalent modification. At present, there is evidence that LPA
3 receptors are phosphorylated in response to agonists and other agents (associated respectively with homologous and heterologous desensitizations)
[46][57]. However, to date, the phosphorylation pattern(s) of this receptor is (are) unknown, which seems to be an important gap in our knowledge.
Studies conducted
in silico showed that the LPA
3 receptor can be phosphorylated by different protein kinases
[57]. Not surprisingly, different isoforms of GRK and PKC are predicted to be responsible for many such phosphorylations; however, other protein kinases such as PKA, PKB/AKT, and some protein tyrosine kinases were present in this
in silico analysis
[97]. Many of these predicted phosphorylation sites could be targeted by several protein kinases
[57][97].
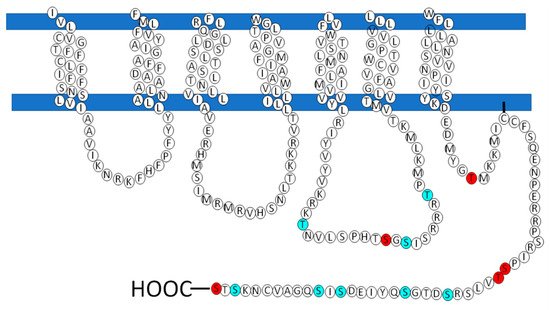
Considering the vital role that GRKs play in homologous desensitization/phosphorylation, the putative sites for the action of this family of kinases on LPA
3 receptor phosphorylation are presented in
Figure 6. These residues were obtained in a new analysis employing different and/or updated software programs, including GPS5 (
http://gps.biocuckoo.cn; Accessed on 3 April 2021), netphorest (
http://netphorest.info; Accessed on 3 April 2021), quokka (
https://quokka.erc; Accessed on 4 April 2021) and NetPhos 3.1 (
http://www.cbs.dtu.dk; Accessed on 4 April 2021). The criterion used to carry out each study was a high threshold. Only residues that were putative targets of GRK, PKA, or PKC and that obtained a high score were considered. Subsequently, we carried out an analysis on the results obtained and chose the residues that were consistently observed in these analyses; these are presented in
Figure 6. The majority of the GRK putative phosphorylation-target residues were found in intracellular loop 3 and the carboxyl terminus region. Not surprisingly, the different software programs used suggested roles of isoforms of the GRK2 and GRK4 subfamilies (
Table 1).
Figure 6. In silico prediction of serine and threonine sites phosphorylated by GRK, PKA and PKC. LPA3 structure is represented, showing (in red) the putative sites targeted by GRK and (in cerulean) putative sites phosphorylated by PKA or PKC.
The possibility that different GRK isoforms may participate in LPA
3 phosphorylation is provocative. It has been proposed that GRK 2 and 3 promote receptor endocytosis by the β-arrestin/clathrin pathway more efficiently than other isoforms. At the same time, GRK 5 and 6 appear to mediate β-arrestin-triggered ERK 1/2 signaling
[98][85][87][93][85][99][100][101][102]. It is important to mention that GRKs, in addition to carrying out GPCR phosphorylation, can phosphorylate other proteins in the cell cytoplasm that are involved in cell signaling, as well as receptor trafficking proteins such as G
αq and G
βγ, PI3K, clathrin, caveolin, MEK, and AKT/PKB, among others
[91][92][103][104][105][106][107][108].
It is noteworthy that the
in silico analysis suggested that PKA and PKC could participate in LPA
3 receptor phosphorylation (
Figure 6 and
Table 2); this result is of interest because it might indicate the involvement of these protein kinases in the heterologous desensitization of this receptor. It has been reported previously that LPA
1–3 receptors can be phosphorylated in response to the pharmacological activation of PCK with phorbol myristate acetate
[57]. However, to the extent of our knowledge, there is no evidence of PKA-induced LPA
3 receptor phosphorylation. It should be noted that the
in silico. Detailed analysis shows a marked overlap between GRK, PKA and PKC, suggesting that some sites could be the target of these clusters of kinases. ( Tabla 1 y Tabla 2 ).
Tabla 2. In silico prediction of residues of LPA3 phosphorylated by PKC and PKA.
| Posición |
Aminoácidos |
PKC / PKA |
| 130 |
S |
PKA |
| 217 |
T |
PKCα / PKCδ / PKCγ |
| 233 |
T |
PKA / PKCδ / PKCι / PKCζ |
| 243 |
T |
PKCi / PKCζ |
| 321 |
S |
PKA / PKCδ / |
| 325 |
S |
PKA / PKC / PKCε |
| 341 |
S |
PKCε |
| 351 |
S |
PKCε |
 +1 point
+1 point