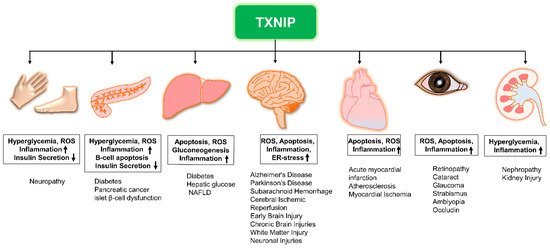
2. TXNIP Is a Potential Therapeutic Target
TXNIP has attracted considerable attention regarding drug development owing to its multiple functions and involvement in metabolic disorders, inflammation, neurodegenerative disorders as well as cancer. Overexpression of TXNIP can be caused by various signals, such as nutritional stimuli, glucose, amino acids, and insulin, suggesting the significance of TXNIP in the regulation of metabolic and neurodegenerative diseases
[83][84][85][86]. By contrast, TXNIP being a participant of apoptosis inducer and metabolic re-programmer works as a tumor suppressor; therefore, downregulation of TXNIP contributes to cancer progression
[87][88][89][90], although such anticancer functions of TXNIP are associated to apoptotic pathways
[91][92]. Thus, TXNIP agonist might help in anticancer treatments, raising yet another debate. In particular, accumulated data provided strong evidence that TXNIP inhibition is a potential therapeutic approach for metabolic disorders and associated diseases
[35][93]. On a cellular level under oxidative-stress the metabolic functions of TXNIP are regulated partially independent of TXN1
[94]. So far, there is no specific inhibitor for TXNIP in clinical trials. Efforts are needed to develop novel TXNIP specific inhibitors to de-intensify the pro-oxidant activities of TXNIP. Although, several in vitro and in vivo studies are underway that either antagonize TXNIP directly or block it through extracellular and intracellular signaling by means of inhibitors, such as small-molecule inhibitors, phytochemicals, and peptides ().
Table 1. Therapeutic modulators of TXNIP. COPD: chronic obstructive pulmonary disease; CTCL: cutaneous T-cell lymphoma; DN: diabetic nephropathy; DR: diabetic retinopathy; HSCs: hematopoietic stem cells; T1DM: type 1 diabetes mellitus; T2DM: type 2 diabetes mellitus.
Several small-molecule drugs have been reported, most of which are being used or under clinical investigation for metabolic and neurological disorders. Verapamil and diltiazem, a nondihydropyridine calcium channel blocker, are used to treat hypertension and angina. It has been observed that verapamil and diltiazem suppress the expression of TXNIP and reverse the β-cell loss in diabetic mice via attenuating TXNIP’s proapoptotic effects
[124][95][98]. Verapamil is in a phase II clinical trial for T1DM, where it is intended to reduce TXNIP expression, increase insulin production, and enhance β-cell mass. Furthermore, the efficacy of verapamil was confirmed in a study in which verapamil administration in diabetic subjects resulted in significantly lower level of fasting serum glucose than in the subjects without verapamil treatment
[97]. Surprisingly, although verapamil shows promising effects in T1DM and at the late stage of T2DM, it does not show any effect in the early stage of T2DM. This may be the reason why verapamil does not reduce TXNIP expression in the liver, muscle, and adipose tissues and, therefore, does not affect the insulin sensitivity of these tissues
[97][125].
Other drugs, such as allopurinol and quercetin, have been found to prevent the overexpression of TXNIP in the rat liver and activation of the NLRP3 inflammasome, and upregulation of sterol-regulatory element–binding protein 1c (SREBP-1c), SREBP-2, liver X receptor α (LXRα), fatty acid synthase, and ROS while downregulating PPARα
[99]. Moreover, several other small-molecule drugs, for example, telmisartan
[126], bakuchiol
[127], vorinostat (SAHA)
[100], trichostatin A (TSA)
[102][128], imatinib
[103], taurine
[104], and troglitazone
[106] can inhibit the expression of TXNIP. Thielen L.A. et al. recently identified a small-molecule inhibitor, SRI-37330, that effectively suppresses TXNIP expression in rats, mice, and human pancreatic islets. In addition, treatment with SRI-37330 reduces glucagon secretion and hepatic glucose production and reverses streptozotocin-induced diabetes
[107]. Nonetheless, further studies are warranted to determine the therapeutic window for clinical trials.
Phytochemicals play a major role in the curative effects of plant-derived products on different diseases, including cancers, autoimmune diseases, and neurological and metabolic disorders. Fisetin and luteolin are natural flavonoids found in vegetables and fruits such as apples, grapes, strawberries, onions, and persimmon. Several in vivo studies have revealed that fisetin treatment of mice downregulates proinflammatory cytokines and ROS production and inactivates TXNIP/MAPK and TLR4/NF-ĸB signaling
[108]. Thus, fisetin exerts beneficial effects on the antioxidant system and diabetes-related diseases as well exhibits anticancer activities and anti-inflammatory properties
[109][110]. Treatment with luteolin protects podocytes from high-glucose induced apoptosis in the mouse podocyte cell 5 (MCP-5) cell line and blocks TXNIP and NLRP3 inflammasome
[129]. Similarly, salidroside suppresses cell proliferation, high-glucose induced oxidative stress, and extracellular-matrix accumulation in rat glomerular mesangial cells (HBZY-1) by inhibiting the TXNIP/NLRP3 signal
[112]. Alkaloids such as cepharanthine and piperine are widely used as antineoplastic, antiallergic, and anti-inflammatory agents and are known to ameliorate diabetic neuropathy
[114][115], whereas piperine stimulates digestive enzymes and lowers lipid peroxidation
[116].
Other phytochemicals have also shown promising effects against different diseases either in vitro or in preclinical models. Among them, metformin
[105], apocynin
[130], curcumin
[131], and ginsenoside (compound K)
[120] exert significant beneficial effects on the antioxidant system, inflammation, cancer, DM, and on many other disorders.
Peptides also contribute to inhibiting TXNIP and are useful for the prevention of several disorders (neurological and metabolic disorders). Thioredoxin-mimetic (TxM) peptides, Ac-Cys-Pro-Cys-amide (CB3), and Ac-Cys-Gly-Pro-Cys-amide (CB4), prevent ROS-related damage by inhibiting p38, MAPK, and c-Jun NH2-terminal kinase (JNK) and by preventing NF-κB nuclear translocation
[121][122]. CB3-treated male leptin-receptor-deficient Zucker diabetic fatty (ZDF) rats show lower inflammation and decreased TXNIP/TBP-2 expression. By contrast, the AMPK pathway is activated, which results in the inhibition of the mTOR-p70S6K pathway. Furthermore, CB3 and CB4 induce apoptosis and reduce caspase 3 cleavage and PARP dissociation in human neuroblastoma SH-SY5Y cells. It has been suggested that these peptides may have a potential to prevent neurological disorders and DM
[122]. Another peptide, TN13, derived from the TXNIP-p38 interaction motif, inhibits the TXNIP–p38 interaction and significantly revives aged hematopoietic stem cells (HSCs). This finding indicates that the interaction between TXNIP and p38 activates the regulatory mechanism of HSC aging and is a possible therapeutic target for the reactivation of aging HSCs
[123].
In recent years, researchers have recognized the role of microRNAs as essential mediators in the control of gene expression via post-transcriptional regulation. Here, we discuss some microRNAs that are potentially relevant for regulating TXNIP and inflammatory diseases (). MiR-20a negatively regulates the NLRP3 inflammatory response in rheumatoid arthritis fibroblast-like synoviocytes. The overexpression of miR-20a reduces TXNIP expression and downregulates the NLRP3 inflammasome and subsequent secretion of cytokine IL-1β, caspase 1, and matrix metalloproteinase 1 (MMP-1)
[132]. Furthermore, the expression of miR-23a is decreased in the blood plasma of patients with central nervous system (CNS) diseases (e.g., ischemic stroke or multiple sclerosis), it also regulates neuropathic pain
[133][134]. Besides, downregulation of miR-23a increases chemokine CXC receptor 4 (CXCR4) expression in a neuropathic pain model
[34].
Table 2. The miRNAs that regulate TXNIP. ALD: alcoholic liver disease; RA FLS: rheumatoid arthritis fibroblast-like synoviocytes.
In addition, several other microRNAs have modulatory functions in the pathogenesis of some diseases. For instance, miR-377 overexpression promotes oxidative stress and increases the production of fibronectin in diabetic neuropathy
[135][136]. Under stress conditions (ERS), the levels of miR-17-5p decrease, leading to inflammasome activation and causing retinal inflammation
[137][138]. In contrast, miR-148a inhibits the expression of TXNIP and prevents the activation of the NLRP3 inflammasome
[78]. MiR-33 increases ROS production and regulates the activity of the NLRP3 inflammasome in chronic inflammatory diseases
[139].
Major efforts are needed to develop drugs that can specifically inhibit TXNIP and are highly effective in overcoming neurological and metabolic abnormalities.
 +1 point
+1 point