The other plastidic FtsH enzymes are believed to be localized in the chloroplast envelope
[19]. Deleting FtsH7 and 9 does not result in any obvious phenotype
[53], and the proteases are not required for PSII repair
[55]. FtsH11 is crucial for growth in long photoperiods
[40] and thermotolerance
[56][57]. FtsH12 was co-immuno-precipitated in a complex with FtsHi1, 2, 4, 5 and NAD-dependent malate dehydrogenase (MDH) and shown to be involved in protein import
[58][59]. In addition to FtsHi1, 2, 4, and 5, even FtsHi3 belongs to the five plastidic FtsH homologues, which are incapable of proteolysis in
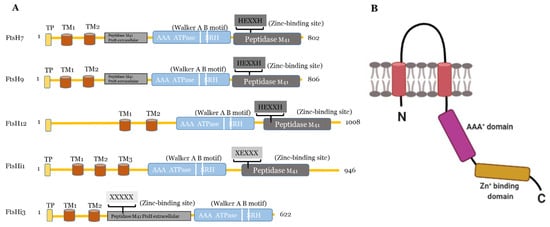
Arabidopsis. The FtsHi enzymes either have a mutation in their HEXXH motif (FtsHi1, 2, 4, and 5), or the entire motif is missing (FtsHi3)
[18][26]. Compared to AtFtsHi1, 2, 4, and 5, FtsHi3 contains a very short C-terminal domain. Interestingly, FtsHi3 also has undergone a domain swap: the whole M41 domain is located at the N-terminal instead of at the C-terminal to the AAA
+ domain
[26]. Comparing the domain organization of AtFtsHi3 with AtFtsHi1, AtFtsH7, 9, 12 (), also AtFtsH7/9 contain this “peptidase M41 FtsH extracellular” domain N-terminal to the AAA
+ domain, which is additional to their protease domain located in the C-terminal to the AAA
+ domain. This additional domain is not present in other FtsHs or FtsHis. Whether the N-terminal “peptidase M41 FtsH extracellular” domain of FtsH7, FtsH9, and FtsHi3 enables these enzymes to form a common complex with a specific function remains to be shown. Three independent pre-protein translocating models (pSSU-TEV-protein A, pL11Flag-TEV-Protein A, pLHCP-TEV-protein A) suggested FtsHi3 to form a 1-MD complex separate from the FtsH12/FtsHi1,2,4,5/MDH complex
[58] and different to the 1-MD TIC complex
[60]. The identity of other components in this complex is unknown.
FTSHi3 is not co-expressed with the tight cluster of
FTSH12/FTSHi1, 2, 4, 5, but instead with a gene encoding OTP51
[26][58][59]. This pentatricopeptide repeat protein is required for the splicing of group IIa introns and impacts photosystem I and II assembly
[61]. The tight co-expression with
FTSHi3 indicates a common function of OTP51 and FtsHi3; therefore, OTP51 is another hypothetical complex partner.
 +1 point
+1 point