1000/1000
Hot
Most Recent
 +1 point
+1 point
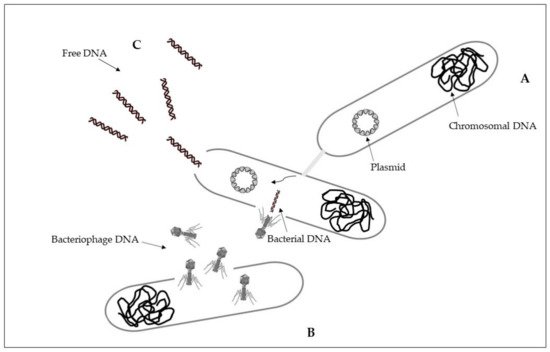
Antibiotic residues originated from aquaculture can select for resistant aquatic bacteria, promoting the spread of antibiotic resistance, even when concentrations were below the minimum inhibitory concentration (MIC) of bacterial strains of the community.
| Country | Mean Percentage (%) | Range Percentage (%) | Year | Isolated Strains | Source |
|---|---|---|---|---|---|
| Bosnia and Herzegovina | 29 | 23–35 | 2016 | 215 | CAESAR |
| Croatia | 29 | 26–32 | 2017 | 1150 | EARS-Net |
| France | 17 | 16–18 | 2017 | 13,328 | EARS-Net |
| Greece | 34 | 32–36 | 2017 | 1464 | EARS-Net |
| Italy | 47 | 46–48 | 2017 | 6945 | EARS-Net |
| Lebanon | 45 | 33–57 | 2016 | 65 | GLASS |
| Portugal | 30 | 29–31 | 2017 | 6424 | EARS-Net |
| Spain | 33 | 32–34 | 2017 | 5557 | EARS-Net |
| Tunisia | 19 | 13–29 | 2017 | 78 | GLASS |
| Turkey | 55 | 53–57 | 2016 | 3670 | CAESAR |
| Country/Area | Isolated Bacteria Species/Genus/Family/Order/Class | Antibiotic-Resistance | Antibiotic-Resistant Genes | References |
|---|---|---|---|---|
| Algeria | Vibrio alginolyticus, V. cholerae, V. fluvialis, V. hollisae | [133] | ||
| Croatia | Aeromonas spp. | [134] | ||
| Eastern Adriatic | Vibrio spp. | flumequinone, chloramphenicol, oxytetracycline | [135] | |
| Egypt | Aeromonas spp. | chloramphenicol, kanamycin, azithromycin | [136] | |
| Egypt | Pseudomonas anguilliseptica | [137] | ||
| Egypt | Aeromonas hydrophila | penicillin, erythromycin | [138] | |
| Egypt | Enterobacteriaceae | cephalosporins, carbapenem | blaKPC (blaCTX-M-15, blaSHV, blaTEM, blaPER-1) | [139] |
| France | Flavobacterium psychrophilum | [140] | ||
| France | Yersinia ruckeri | [141] | ||
| Greece | Acinetobacter spp., Bacillus spp., Pseudomonas spp., Staphylococcus spp., Stenotrophomonas spp. | tetracycline | tetA, tetK, tetC, tetE, tetM | [127] |
| Italy | Enterococci | ampicillin, gentamicin | tetM, tetL, tetO, ermB, mef | [142] |
| Italy | Aeromonas spp., Photobacterium spp., Shewanella spp., Vibrio spp. | tetracycline, flumequine, trimethoprim | [122] | |
| Italy | Photobacterium damselae ssp. piscicida, Vibrio fluvialis, V. alginolyticus, V. parahaemolyticus, V. metschnikovii | ampicillin, carbenicillin, kanamycin, cefalothin | [143] | |
| Italy | Aeromonas spp. | ampicillin, amoxicillin, cephalothin, erythromycin, streptomycin, sulfadiazine, trimethoprim | [125] | |
| Italy | Shewanella algae, Vibrio spp. | beta-lactams, quinolones, tetracyclines, macrolides, polymyxins, chloramphenicol, fosfomycin, erythromycin | blaOXA-55-like, blaAmpC, mexB-OprM, mdtG, mdlB,tet34, tet35, tetR, eptA, cat, mdtL |
[46] |
| Lebanon | Streptococcus pneumoniae | polymyxins, chloramphenicol, fosfomycin, erythromycin | [144] | |
| Spain | Aeromonas spp., Salmonella spp., Vibrio mimicus, V. furnissii. | oxytetracycline, nitrofurantoin, oxacillin, sulfomethoxazole/trimethoprim | [145] | |
| Spain | Flavobacterium psychrophilum | oxytetracycline, florfenicol | [146] | |
| Spain | Aeromonas salmonicida | nalidixic acid, oxytetracycline | [147] | |
| Tunisia | Vibrio alginolyticus | ampicillin, erythromycin, kanamycin, cefataxime, streptomycin, trimetoprim | [148] | |
| Tunisia | Escherichia coli | tertracycline, streptomycin, ampicillin, trimethoprim, sulfamethoxazole | tetA-tetB | [149] |
| Turkey | E. coli, coliforms, fish pathogens | sulfamethoxazole, ampicillin, sulfamethoxazole, imipenem, aztreonam | ampC, blaCTX-M1, tetA, sul2, blaTEM | [150] |
| Turkey | Y. ruckeri | floR, sulI, tetC, tetD, tetE | [151] | |
| Turkey | Y. ruckeri | erythromycin, florfenicol, sulfonamide, tetracycline, trimetophrin | ermB, ermY, floR, su/I, suffll, tetA-tetG | [152] |
| Turkey | Aeromonas media, A. rivipollensis, A. salmonicida, Bacillus pumilus, B. zhangzhouensis, Hafnia alvei, Kluyvera intermedia, Pantoea spp., Pseudomonas spp., P. protegens, Staphylococcus spp., Gammaproteobacteria, Betaproteobacteria, Enterobacteriales, Burkholderiales | sulfamethacin, sulfamerazine, erythromycin, tetracycline | [153] |





