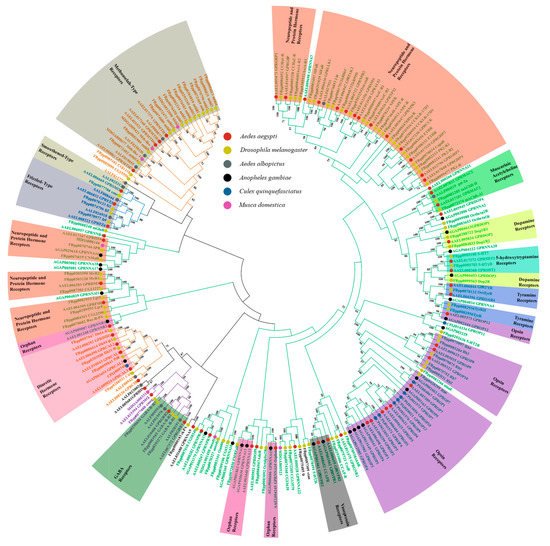
3.1. Brain Tissue and Central Nervous System
Several GPCRs have been identified as being predominately expressed in the brain and CNS, corresponding to neuropeptide expression in diverse insect species. A neuropeptide bona fide natalisin receptor found to be highly expressed in the CNS of
D. melanogaster,
T. castaneum and
B. mori regulates sexual activity and fecundity in insects
[20]. A mutant
methuselah gene expressed in the brain of
Drosophila, a model insect often used to investigate gene functions, plays a critical function in oxidative stress resistance
[136]; a dopamine/ecdysteroid receptor that is a head-specific expression gene is also overexpressed in
Drosophila adults and embryos
[97]. Another study also found the dopamine/ecdysteroid receptor (
DopEcR) to be highly expressed in
Drosophila nervous system and brain tissues, and its mutant has been implicated in the insect’s resistance to alcohol
[68]. A
Drosophila sex/myp-inhibiting peptide receptor expressed in the CNS is highly conserved in insects
[19], while an octopamine receptor
DmOctα2R transcript present at much higher levels in
Drosophila males than in females is mainly expressed in the head of adults
[138] and a
Drosophila neuropeptide capa GPCR expressed in larvae central nerve system is responsible for sensitivity to desiccation stress
[37]. Of two lGRs (
LGR3/LGR4) characterized in diverse
Drosophila tissues,
Lgr3 transcripts were predominately expressed in the CNS, while
Lgr4 transcripts were highly expressed in the CNS of the wandering larval stage. In adults, the
Lgr4 transcripts showed significantly high expression in the male thoracic-abdominal ganglion and brain tissues
[139].
The silkworm,
B. mori, which is classed as a beneficial insect, is another model species frequently used in gene functional studies. A neuropeptide GPCR A4 receptor gene (
BomNPFR) amplified from its brain tissue was predicted to be involved in food intake and growth processes
[130]. Three GPCR receptors responding to neuron ion transport peptides (ITPs) were identified in silkworm larvae using tissue specific expression, revealing that bngr-A2 was mainly expressed in the brain, with no expression in the CNS, and bngr-A34 was not present in nervous tissues
[21]. A neuropeptide allatotropin receptor was significantly overexpressed in the corpora cardiaca and brain of
B. mori, indicating the potential involvement of juvenile hormone (JH) biosynthesis processes
[140]. In honeybees, an important beneficial insect that plays a critical role in pollination, an octopamine receptor was identified in the honeybee brain
[63]. A tyramine receptor was later identified that was highly expressed in nurse bees and forager brain neuropils
[141].
The red import fire ant,
Solenopsis invicta, is an invasive urban insect species whose gene expression and functions have been widely studied over the last couple of decades. Ten GPCR gene expressions have exhibited significant differences in the brain tissue of workers, alate virgin queens, and mated queens
[90]. One neuropeptide F-Like receptor was isolated from
S. invicta and its differential expression levels in the brain suggest its potential function to be feeding regulation for mated queens
[48]. In the medically important kissing bug,
Rhodnius prolixus, an important Chagas disease vector, GPCRs have been found to be involved in multiple physiological pathways. Three variants of a pyrokinin-1 receptor gene characterized in different tissues of fifth instar larvae of
R. prolixus revealed two variants that were mainly expressed in the CNS
[142]. A corazonin receptor was found to be overexpressed in the brain and involved in
R. prolixus heartbeat control
[143] and a serotonin type 2b receptor transcript has also been shown to be upregulated in the CNS of
R. prolixus [144]. To predict the potential function of a kinin receptor in
R. prolixus blood post-feeding, a kinin receptor was characterized in various tissues and found to be overexpressed in the CNS of late instar larvae
[56]. In mosquitoes, which are responsible for transmitting a number of human and animal diseases, GPCRs have been identified as being involved in mosquito blood feeding and insecticide resistance. The overexpression of an allatotropin GPCR receptor (
AeATr) gene was characterized in the nervous system and corpora alata-corpara cardiac complex of
Ae. aegypti. Blood feeding depressed the transcript level of
AeATr, and was associated with JH biosynthesis in mosquitoes
[32]. A rhodopsin-like GPCR overexpressed in the head of adult mosquitoes was found to play a critical role in the development of permethrin insecticide resistance in the mosquito,
Cx. Quinquefasciatus [43]. Tissue specific studies conducted on this GPCR gene revealed its significantly high expression in the insect’s brain tissue along with a G-protein alpha subunit, two adenylyl cyclase and one protein kinase A gene, playing important roles in the GPCR-leading intracellular pathway in the insect’s neuron system and regulating its insecticide resistance
[44]. A corazonin neuropeptide receptor (
MdCrz) has been found to be overexpressed in the larval CNS of the house fly, another medically important pest, including in its ventral nerve cord, the protocerebral DL neurons of the brain lobes and its
vCrz neurons. Although it was overexpressed in the heads of both male and female houseflies, there was no expression in other body tissues, indicating that
MdCrz is a brain-specific expression gene
[145].
The red flour beetle,
T. castaneum, is a stored product pest that causes problem for communities around the world. The spatial expression of a 5-HT7-type serotonin receptor identified in
T. castaneum showed the highest expression levels to be in the head of both male and female adults, predominantly expressed in the brain but accompanied by high levels of expression in the optic lobes, predicting the functional importance of this receptor in neural processes
[134]. A D2-like dopamine receptor gene predominantly expressed in the head and CNS of
T. castnaeum adults
[146] and an inotocin receptor was found to be overexpressed in the head of the insect’s early larval stage
[147]. In the desert locust,
S. gregaria, an agricultural pest species, two novel octopamine receptors were characterized in adults, with
SgOctαR being overexpressed in the CNS, including the brain, optic lobes, subesophageal ganglion and thoracic ganglions, while
SgOctβR was overexpressed in the CNS. Interestingly, these receptors were highly expressed in the long-term gregarious locusts but not in the solitarious locusts, suggesting the receptors’ function in locust behavior
[148]. GPCRs have also been widely studied in other agricultural pest species. The GPCR
NIA42 was found to be highly expressed in the brain and abdominal integuments of the adult brown planthopper,
N. lugens, where it was linked to the neuropeptide NI-elevenin that is involved in the regulation of planthopper melanization
[70][149]. A novel octopamine receptor gene expression has been characterized in diverse tissues of the fifth-instar larva rice stem borer,
Chilo suppressalis, showing a high level of expression in the nerve cord
[150]. A large screen of GPCRs that was characterized in
C. suppressalis identified 51 putative GPCR genes. The expression of these genes was examined in tissues of the insect’s CNS, fat body, gut and hemocytes, with most of the receptor genes being highly expressed in the CNS
[151]. A serotonin receptor (
Pr5-HT8) was identified as being highly expressed in the nerve cord of the larva small white butterfly,
Pieris rapae. 5-hydroxytryptamine (5-HT) has been identified as a neurotransmitter that plays critical roles in the regulation of a number of physiological processes, which is consistent with the expression of
Pr5-HT8 in the butterfly’s CNS
[133]. An allatotropin receptor has also been found to be overexpressed in the brain, thoracic ganglion and abdomen ganglion of the bollworm,
H. armigera [152].
3.2. Digestion and Reproduction Systems
In mosquito species, GPCRs are known to be expressed in the digestion system and involved in blood-feeding behavior. A leucokinin receptor has diverse functions, responding to multiple kinins in the mosquito,
Ae. aegypti, and is expressed in the mosquito’s hindgut and Malpighian tubules
[153]. Piermarini’s group also identified several GPCRs and metabolic genes upregulated or downregulated in the Malpighian tubules of blood-taken
Aedes albopictus [154]. An allatotropin GPCR receptor (
AeATr) gene has also been found to be overexpressed in the ovary of adult mosquitoes
[32]. In another blood-feeding insect,
R. prolixus, a serotonin type 2b receptor transcript was overexpressed in the Malpighian tubules, salivary glands and guts
[144]. A kinin receptor has also been found to be highly overexpressed in the gut system of late instar larvae of
R. prolixus [56]. Two variants of a pyrokinin-1 receptor gene were mainly expressed in the male testes and prothoracic glands of fifth instar larvae of
R. prolixus [142]. In
Drosophila, several GPCRs were identified as overexpressed in the digestion and reproduction systems. These
Drosophila Lgr4 transcripts were highly expressed in the gut system of the wandering larval stage, and significantly highly expressed in the male midgut and crop
[139]. Additionally, a sex/myp-inhibiting peptide receptor was highly expressed in the male
Drosophila reproduction organs
[19] and an octopamine receptor gene was highly expressed in the Malpighian tubules, with lesser amounts found in the midgut and hemocytes of the fifth-instar larva rice stem borer,
C. suppressalis [150]. Several GPCRs in
C. suppressalis were overexpressed in the gut system
[151]. A neuropeptide F-Like receptor detected in the gut and reproduction systems of
S. invicta suggests its potential function was involved in the feeding regulation of mated queens
[48]. One ITP gene identified in
B. mori, bngr-A2, was found to be mainly expressed in the reproduction system
[21]. A serotonin receptor (
Pr5-HT8) is known to be highly expressed in the Malpighian tubules, fat body and midgut of larvae of the small white butterfly,
P. rapae [133].
3.3. Other Insect Organs
In addition to the GPCRs identified in the CNS, digestive and reproductive systems in insect species, many GPCRs have also been characterized in other organs. In
Drosophila adults, Lgr3 transcripts were found to be highly expressed in the female fat body and uterus, and the male salivary glands
[139]. An adipokinetic hormone receptor was predominately expressed in the fat body of the oriental fruit fly,
B. dorsalis, and was also involved in its triacylglycerol mobilization and sexual behavior
[62]. Forty six putative GPCR transcripts have been isolated from the foreleg tissues of the cattle tick,
Phipicephalus australis, providing valuable information for GPCR studies of signal transduction, host preference and mating behavior in insects
[155]. An allatotropin receptor has been identified that is overexpressed in the male bumblebee accessory glands, predicting its potential involvement in JH biosynthesis
[156] and in
B. mori a sex peptide receptor has been found to be highly expressed in the prothoracic gland, predicting the critical roles of receptor in regulating ecdysteroidogenesis
[28]. A novel octopamine receptor,
SgOctβR, was also overexpressed in the flight muscles of
S. gregaria, suggesting its involvement in locust gregarious behavior
[148], while a GPCR receptor,
NlA42, was significantly highly expressed in the integument and salivary gland of the brown planthopper indicating its function in the insect’s melanization
[149].
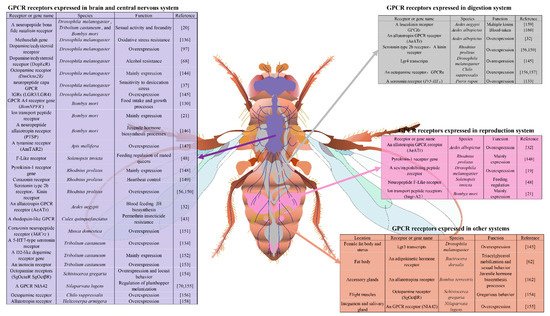
Figure 2. Tissue specific expression of GPCR genes in insect species. Insect GPCRs are mainly expressed in the brain and central nervous system, highlighted in light blue; those in the digestive system are highlighted in gray; those in the reproductive system are highlighted win light purple; those in other organs are highlighted in orange. Twenty-six GPCRs that are highly expressed in the brain and central nervous system in 13 insect species are involved in sexual activity and fecundity, oxidative stress resistance, desiccation stress, food intake and growth, juvenile hormone biosynthesis, feeding regulation, heartbeat control, permethrin insecticide resistance, behavior and cuticle melanization.
However, while the precise functions of the GPCRs in specific tissues are still in need of further characterization, the impact of the GPCRs on insect behaviors can now be predicted to some extent based on the functions of those specific tissues in insects. The importance of GPCRs in insects can be partially appreciated by considering their distribution, location and function within the cells of specific tissues. In humans, if any of the fundamental processes governed by GPCRs in specific tissues are dysfunctional, this will lead to acute or chronic diseases
[157][158][159][160]. The physical location and expression of GPCRs may provide a direct mechanism for the transduction of extracellular messages into tissue responses and functions. Consequently, the identification of the GPCRs in brains and CNS of insects could shed new light on the important roles they play in neuronal firing, the regulation of ion transportation across cell membranes and the modulation of neuron membrane potentials in other species.
 +1 point
+1 point