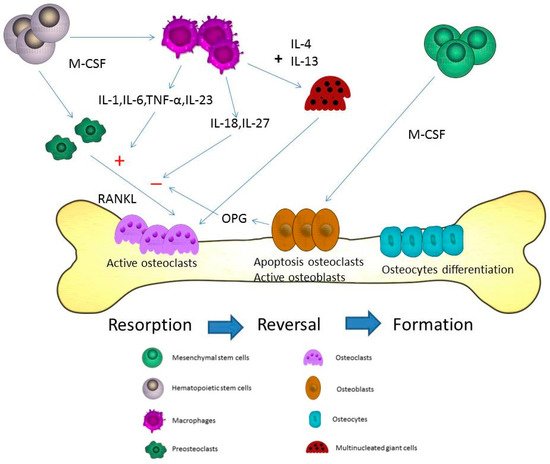
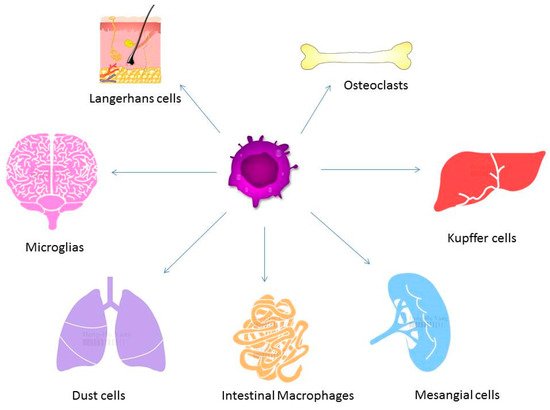
Macrophages play a major role in the innate and adaptive immune system. Differentiation of macrophages can be found in various tissues including liver, lung, brain, and bone marrow. Macrophages are divided into M1, which express proinflammatory mediators, and M2 that are involved in anti-inflammatory reactions
[21]. In the innate immune system, macrophages can execute phagocytosis and opsonization
[22]. Different cell receptors of CD14, Fcγ, and CD25 are found in macrophages. Major histocompatibility complex class II molecules and CD23 are also found. These receptors are important for the progression of phagocytosis. IL-4 and IFN-γ can regulate the different functions of macrophages. Macrophages, in turn, may be activated to induce IL-6, TNF-α, IFN-γ, complement protein, and prostaglandins in the immune system. The cytokines expressed by macrophages associated with stimulation or inhibition of osteoclastogenesis include IL-6, IL-18, IL-23, IL-27, and TNF-α
[23]. IL-6, a proinflammatory cytokine, can activate osteoclastogenesis
[24]. During the inflammation state, proinflammatory cytokines including TNFα, IL-1β, and IL-6, may promote the differentiation and activation of osteoclasts
[25]. IL-18 secreted via macrophages may regulate the Th1 differentiation and the IFN-γ production, and is an inhibitor of the TNF-α mediated osteoclastogenesis
[26]. Among osteoporotic women, decreased levels of serum IL-18 binding protein and elevated levels of serum IL-18 are observed
[27][28]. IFN-γ has a dual role in osteoclasts including the promotion of osteoblast differentiation and inhibition of bone marrow adipocyte formation in different stages
[29]. IFN-γ can activate macrophages, but macrophages can secrete IL-18 to regulate the IFN-γ production. IL-23 has been shown to activate osteoclasts
[30]. Adding IL-23 to bone marrow stromal cells led to an increased differentiation towards the osteoblast lineage
[31]. In the femur of a rat osteoporosis model, IL-23 is reduced after adequate estrogen therapy for improvement of bone mineral density
[32]. IL-27 may suppress the expression of RANKL in Th17 cells and CD4+ T cells
[33]. IL-27 also inhibited osteoblast apoptosis through increased Egr-2 expression
[34]. TNF-α from macrophages may induce indirect osteoclastic activation through RANKL in bone remodeling
[35]. Therefore, inflammatory arthritis, such as rheumatoid arthritis, can induce progressive bone loss when the disease is poorly controlled. The different cytokines of macrophages associated with osteoporosis are shown in .
 +1 point
+1 point