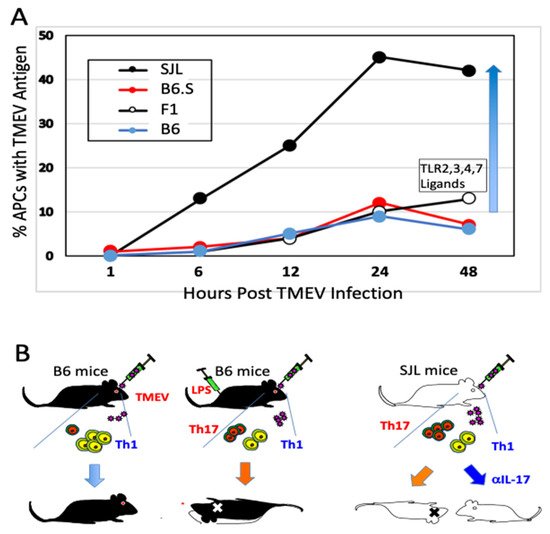
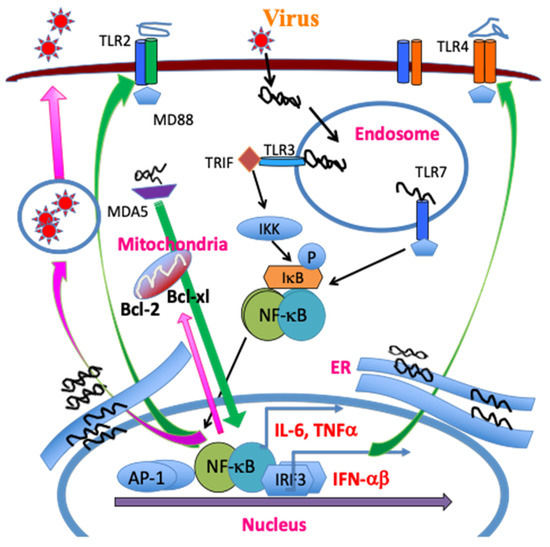
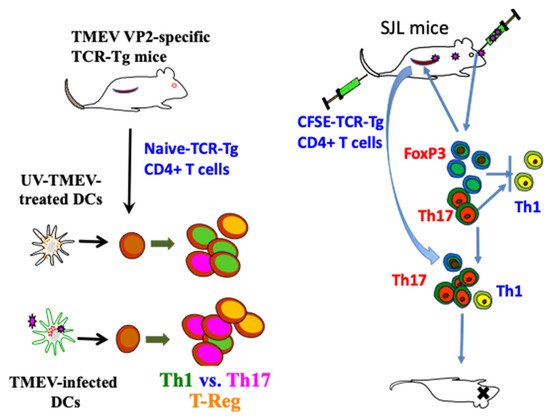
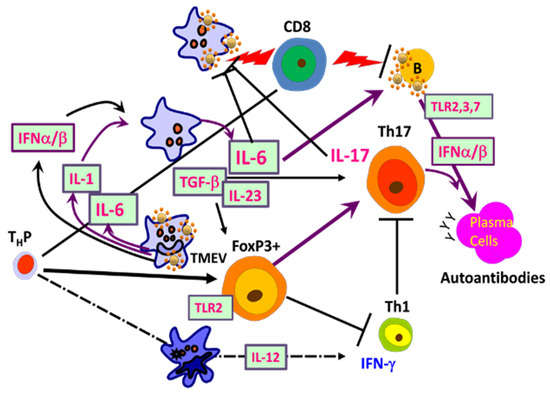
As demyelination is closely linked to viral persistence
[159][160][161], TMEV-specific cytotoxic CD8
+ T cells producing IFN-γ and perforin (Tc1) are likely to play an important role in protection and/or resistance
[162][163][164]. TMEV-specific cytotoxic CD8
+ T lymphocytes (CTL) appear to damage virus-infected, myelin-producing oligodendrocytes and other cell types
[165][166][167][168]. Many investigations further confirmed the role of Tc1 cells by antibody-mediated CD8
+ T cell depletion
[169], and using Class I deficient mice
[170][171][172]. Rodriguez and his colleagues proposed that CD8
+ T cells are necessary for the manifestation of clinical symptoms using the DA strain of TMEV
[163][167]. However, β
2M-deficient or perforin-deficient mice on a resistant background are susceptible to both demyelination and clinical disease
[164][170][171][172]. Furthermore, β
2M-deficient mice with the susceptible SJL background displayed similar exacerbation of TMEV-IDD
[173]. These results indicated a protective role of CD8
+ T cells in the development of TMEV-induced demyelinating disease in both resistant and susceptible mice. Moreover, the presence of a high level of CTL in resistant mice and a low level in susceptible mice
[174], and the resistance to TMEV-IDD in susceptible mice adoptively received CD8
+ T cells
[175], further support the protective function of CTL. In resistant B6 mice, the majority (50% to 70%) of CNS-infiltrating CD8
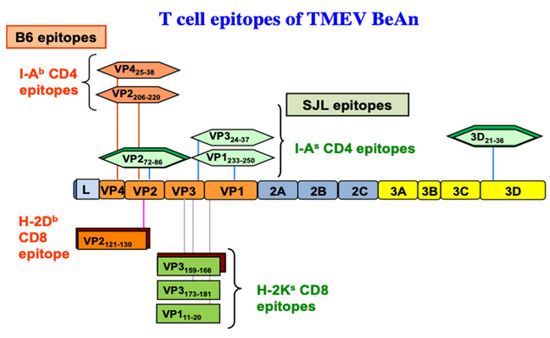
+ T cells recognize VP2
121–130 [176][177], and two minor populations (<10%) react with VP2
165–173 and VP3
110–120 capsid epitopes
[178] based on the production of IFN-γ (). Similarly, CNS-infiltrating CD8
+ T cells of virus-infected SJL mice react with one predominant (VP3
159–166,) and two subdominant capsid epitopes (VP3
173–181, and VP1
11–20)
[35]. During the early stages of viral infection, a lower level of virus-specific CD8
+ T cells in SJL mice was observed
[178]. In addition, the resistance of (B6xSJL)F1 mice is associated with a higher level of the initial virus-specific H-2b-restricted CD8
+ T cell responses compared to the H-2s-restricted CD8
+ T cell responses
[37]. These results further suggest that Tc1 cells play an important protective role in preventing TMEV-IDD by clearing viral loads from the CNS. There is, however, a possibility that certain CD8
+ T cell populations play a pathogenic role, perhaps depending on epitope-reactivity or cytokine production, in TMEV-induced demyelination
[165][166][167][179]. Similar CTL-mediated immunopathology was reported with the lymphochoriomeningitis virus (LCMV) and Coxsackie B virus in mice
[180][181][182].
 +1 point
+1 point